2. INGESTIÓN,
DIGESTIÓN Y EXCRECIÓN. Fagocitosis y pinocitosis.
3. PASO DE
SUSTANCIAS A TRAVÉS DE LA MEMBRANA. Transporte pasivo: ósmosis, difusión
simple (iones y gases) y difusión facilitada. Transporte activo.
1. Nutrición
autótrofa y heterótrofa
Como
veíamos en el tema 7, llamamos nutrición al intercambio material y
energético entre el ser vivo (en este caso, la célula) y el medio ambiente.
Este intercambio no se reduce a la captación de nutrientes y a su posterior
degradación, como muchas veces se hace en el lenguaje vulgar. Junto a la ingestión
y digestión de alimentos, abarca los procesos de circulación de
nutrientes, excreción de productos de desecho y metabolismo en
general (aunque dentro de éste haremos especial mención de la fotosíntesis, la
fermentación y la respiración celular).
El
proceso de la nutrición propiamente dicho tiene lugar en el interior de las
células, pues en ellas se desarrollan las reacciones químicas que transforman
los alimentos en sustancias propias, los degradan para obtener energía de este
proceso, y se eliminan los productos resultantes de esa degradación. Pero a
veces es necesaria una degradación previa de los alimentos para que éstos
puedan incorporarse a las células. Es lo que denominamos digestión
extracelular, que estudiaremos en el nivel orgánico.
Las
sustancias nutritivas deben estar en condiciones para atravesar las membranas
celulares. Y las membranas celulares sólo pueden ser atravesadas por los gases,
el agua y las sustancias disueltas (siempre que sean sencillas). Esto último
hace que sea preciso realizar en la célula lo que denominamos digestión
intracelular, que no es sino el desdoblamiento de moléculas complejas en
moléculas sencillas.
Como
sustancias nutrientes deben considerarse no sólo el agua y los alimentos
propiamente dichos (orgánicos y sales minerales), sino también los gases como
el oxígeno (en animales y en vegetales) y el CO2 (en vegetales), y
la energía luminosa o química del medio circundante.
Según
cuál sea la naturaleza de las sustancias nutritivas que toman los seres vivos y
la fuente de energía, distinguimos dos tipos de nutrición: autótrofa y heterótrofa.
a)
Nutrición autótrofa
Se
lleva a cabo con materiales inorgánicos: agua, CO2, y sales
minerales disueltas. Al tratarse de sustancias con muy poca energía, los seres
que presentan nutrición autótrofa deben tomar esa energía de la luz solar o de
determinadas reacciones químicas exotérmicas que se producen en el medio
ambiente. Con esa energía y a partir de la materia inorgánica, los seres
autótrofos sintetizan su propia materia orgánica: son los únicos seres vivos
capaces de sintetizar materia orgánica a partir de materia inorgánica.
Dependiendo
de la fuente de energía utilizada distinguimos dos tipos de nutrición
autótrofa: fotosíntesis y quimiosíntesis.
-En
la fotosíntesis (propia de los organismos fotosintéticos o fotolitótrofos, como los denominamos en el tema 7) la
fuente de energía en la luz solar (son las plantas, las algas, las
bacterias rojas del azufre y las bacterias verdes del azufre).
-En
la quimiosíntesis (propia de los quimiosintéticos o quimiolitótrofos)
la energía se obtiene de reacciones oxidativas exotérmicas (bacterias
del hidrógeno, bacterias incoloras del azufre, bacterias nitrificantes
y bacterias férricas).
b)
Nutrición heterótrofa
Se
realiza con materiales orgánicos de elevado contenido energético (glúcidos, lípidos y prótidos). Aunque los organismos heterótrofos
también incorporan agua y sales minerales, son incapaces de aprovechar la
energía solar o la que se desprende de las reacciones químicas oxidativas de
moléculas inorgánicas. Por este motivo todo ser heterótrofo debe vivir a
expensas de los autótrofos o sobre materia orgánica en descomposición.
Son
heterótrofos los animales, los hongos, los protozoos y la mayoría de las
bacterias.
2. Ingestión,
digestión y excreción
a)
Ingestión
Si
exceptuamos los gases las sales minerales y algunas moléculas sencillas como la
glucosa, la mayor parte de los nutrientes tienen que sufrir un proceso de
desintegración antes de ser absorbidos en forma de sustancias más sencillas por
la célula. Para esto pueden pasar por lo que denominamos digestión
extracelular, tras la cual se obtienen monosacáridos, glicerina, ácidos
grasos o aminoácidos que pueden atravesar las membranas de las células.
En
otros casos los nutrientes complejos son introducidos en la célula por un
proceso de endocitosis y allí padecen la digestión intracelular.
Existen dos modalidades de endocitosis: fagocitosis y pinocitosis.
-Fagocitosis.
Protozoos de tipo ameba u otros, al tener al alcance un alimento emiten pseudópodos que rodean la partícula.
Así
originan una cavidad llamada vacuola digestiva. Posteriormente se
vierten en ella enzimas digestivas que realizan la digestión intracelular. Los
productos resultantes pasarán al citoplasma a través de la membrana de la
vacuola.
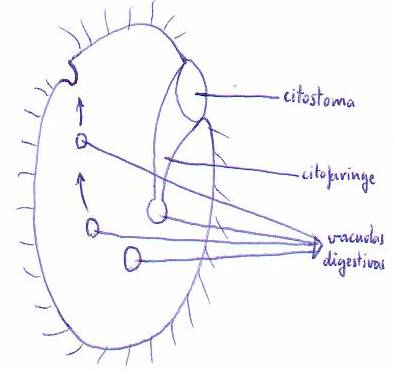
Otros
protozoos -como los ciliados- cuya membrana no permite la formación de pseudópodos, presentan una abertura llamada citostoma seguida de un conducto o citofaringe.
En el extremo
de la citofaringe se forman las vacuolas digestivas.
En
metazoos la fagocitosis se utiliza más como mecanismo de defensa que como
proceso nutritivo.
-Pinocitosis. Es la
ingestión de líquidos por formación de invaginaciones o bolsas de la membrana
celular que se estrangulan y dan lugar a vacuolas digestivas. Se da en
organismos unicelulares y en diversas células de pluricelulares
(particularmente en las que tapizan las cavidades digestivas).
b)
Digestión
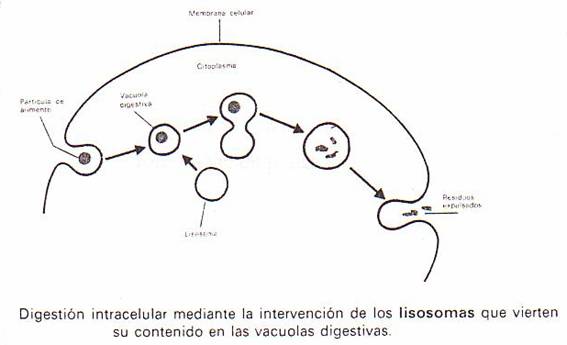
La
captura de sustancias por los procesos de fagocitosis y pinocitosis
no supone su digestión. Se requiere la presencia de lisosomas que vierten sus
enzimas hidrolíticos en la vacuola, al fusionarse con
ésta.
Las
vacuolas digestivas tienen en su primera fase una reacción ácida, lo cual
provoca la muerte de la presa (si se trataba de un microorganismo) y facilita
el comienzo de la digestión. A continuación la vacuola toma contacto y se
fusiona con un lisosoma: cambia entonces su pH que se
vuelve alcalino, y recibe del lisosoma las enzimas hidrolíticas
que provocan la digestión. (Recordemos que llamábamos lisosomas primarios
a los que todavía no se habían fusionado con la vacuola digestiva, y lisosomas
secundarios al fruto de la unión de lisosomas primarios con las vacuolas de
fagocitosis).
A
medida que avanza la digestión los productos resultantes atraviesan la membrana
de la vacuola (mejor dicho, del lisosoma secundario) y se incorporan al
citoplasma.
c)
Excreción
Los
residuos no digeribles son expulsados al exterior por un proceso de exocitosis en el que la vacuola se fusiona con la
membrana exterior y vierte su contenido. En algunos protozoos, como los
Ciliados, existe en la membrana un poro de expulsión (citopigio)
por el que la vacuola digestiva realiza siempre la egestión.
3. Paso de
sustancias a través de la membrana (transportes pasivo y activo)
Las
células captan del medio ambiente en el que viven los nutrientes necesarios
para su vida, y a la vez eliminan los productos de desecho de su metabolismo.
Este intercambio bidireccional de sustancias se
realiza gracias a las propiedades de la membrana plasmática quedelimita la célula.
La
membrana presenta una permeabilidad selectiva, es decir, permite el paso de
algunas sustancias mientras que impide el de otras. Y el paso de las primeras
lo permite con una dificultad variable.
Se
trata de una membrana semipermeable que, como hemos visto para este tipo
de membranas, permite que en la célula se desarrollen fenómenos osmóticos. Pero
sólo es rigurosamente semipermeable en casos excepcionales, ya que también debe
captar diversos nutrientes (no sólo agua).
Por
lo tanto, la membrana plasmática independiza relativamente a las células del
medio líquido extracelular que las rodea, y su actividad permite que la
composición del medio líquido intracelular y extracelular sean muy diferentes:
algo imprescindible para la vida.
líquidolíquido
extracelularintracelular
Na+142 meq/l10 meq/l
K+5 meq/l141 meq/l
Ca2+5 meq/l<1 meq/l
Mg2+3 meq/l58 meq/l
Cl-103 meq/l4 meq/l
HCO3-28 meq/l10 meq/l
fosfatos4 meq/l75 meq/l
SO4=1 meq/l2 meq/l
a)
Transporte pasivo
Denominamos
transporte pasivo al que se da a través de la membrana a favor de gradiente y
por lo tanto sin gasto de energía.
1) Ósmosis. La membrana
plasmática se comporta como una membrana semipermeable. Por eso resulta lógico
que el intercambio hídrico (de agua) entre la célula y el medio que la rodea
está regulado por las leyes de ósmosis.
La
membrana semipermeable permite el paso del agua -pero no el de solutos- de un
sitio a otro hasta que se igualan las concentraciones a ambos lados.
Cuando
la célula se halla rodeada de una solución hipotónica respecto a la de su
interior, penetra agua en la célula. Si la solución extracelular es
hipertónica, sale agua de la célula.
El
fenómeno de la ósmosis explica satisfactoriamente el transporte pasivo de agua.
2) Difusión simple. En
términos físicos difusión es el proceso por el cual un gas o una sustancia en
disolución, debido al movimiento térmico al azar de las partículas de la
solución (movimiento Browniano) tiende a ocupar todo el volumen disponible.
Si
ponemos en contacto dos soluciones con diferente concentración de una misma
sustancia, hay un movimiento neto por la difusión de la que tiene concentración
alta a la de baja concentración.
La
velocidad de la difusión es directamente proporcional a:
-la
diferencia de concentraciones,
-el
área de la superficie a través de la cual se difunde la sustancia,
-la
temperatura;
e inversamente
proporcional a:
-el
radio de la molécula,
-la
distancia que debe recorrer.
En
la difusión simple de sustancias a través de la membrana hay que distinguir 2 formas
principales: la de sustancias lipófilas a través de
la fase lipídica; y la de sustancias
hidrófilas de pequeño tamaño a través de los poros de la membrana. En
cualquiera de los 2 casos el transporte por simple difusión se verifica según
las leyes físicas de la difusión, ósmosis, equilibrio Donann,
y siempre a favor de gradiente de concentración.
*Difusión
de iones. Además hay que tener en cuenta que la difusión de
partículas cargadas (como los iones) depende no sólo del gradiente de
concentración (químico), sino también del gradiente de cargas
(eléctrico) a través de la membrana plasmática, es decir, que los iones
(cargados eléctricamente) tienden a difundirse a favor de gradiente
electroquímico.
El líquido que
baña las células del organismo contiene grandes cantidades de iones difusibles como el Na+
y el Cl-, mientras que en el interior hay muchas sustancias que se
ionizan desprendiendo cationes metálicos como el Na+
y grandes aniones orgánicos (que llamaremos R-), los cuales, por su
gran tamaño no pueden atravesar la membrana.
Cabría
esperar:
célulamedio externo
inicial:
[Na+] = 7'5[Na+] = 7'5
[R-]
= 7'5[Cl-]
= 75
equilibrio:
[Na+] = 10[Na+] = 5'0
[R-]
= 7'5[Cl-]
= 5'0
[Cl-]
= 2'5
pues
la tendencia es a alcanzar el llamado equilibrio de Donann
en el que [Na+] x [Cl-] en la
célula es igual a [Na+] x [Cl-]
en el medio externo.
Y
la membrana sólo permite el paso de Cl- y de Na+
pero no de los grandes aniones que hay dentro de la célula. Según esto Na+ y Cl- quedarían desigualmente
repartidos en ambos lados de la membrana y se daría a la vez un estado de
neutralidad iónica.
Sin
embargo, en las células vivas se observa (por microelectrodos)
que en el interior existe un predominio de cargas negativas y en el exterior de
positivas. Es el llamado potencial de membrana.
Este
potencial se genera debido a que la membrana presenta, además de la difusión
iónica, otro mecanismo que altera el equilibrio eléctrico: la bomba de Na+ y K+.
Si
estudiamos la tabla reflejada al comienzo de este tema, vemos que los iones Na+ y K+ no
difunden libremente. Hay una baja concentración de Na+
en el interior de la célula por un bombeo hacia el exterior de esos iones
mediante un mecanismo que veremos en el transporte activo. Así se explica la
electronegatividad del interior celular.
Por
el contrario, los K+ son introducidos. Eso
sí, por un mecanismo semejante.
Cuando
una membrana mantiene esta situación se dice que está polarizada.
El
mecanismo de transporte de Na+ y K+ a través de la membrana está coordinado. Por
eso a la bomba iónica se la conoce como bomba de Na+-K+. Tiene gran importancia, pues, como
veremos, interviene en la transmisión del impulso nervioso y en la contracción
muscular.
*Difusión
de gases. Se realiza también a favor de gradiente de
concentración, de acuerdo con las leyes físicas de la difusión: de la zona de
mayor tensión o presión parcial a la de menor.
Este
mecanismo actúa en el intercambio de O2 y de CO2 dentro
del sistema circulatorio. Esos gases no requieren poros: pasan a través de la
fase lipídica por ser solubles en lípidos.
3) Difusión facilitada.
DANIELLI, en 1954 llamó difusión facilitada al fenómeno por el que algunas
sustancias presentaban una difusión a través de las biomembranas
mayor que la que cabría esperar por su solubilidad lipídica
o por su magnitud molecular, aunque su paso se verificaba siempre a favor de un
gradiente químico de concentración (es decir, sin consumo de energía
metabólica).
Igual
que en la difusión simple, el transporte se desarrolla a favor de un gradiente
de concentración. Pero, a diferencia de la difusión simple, en la difusión
facilitada hay una interacción de la sustancia transportada con la membrana y
el mecanismo es saturable: a pequeñas concentraciones hay una relación lineal
entre la concentración de soluto y la velocidad de paso, pero cuando la
concentración de soluto aumenta indefinidamente llega un momento en que la
velocidad de transporte no aumenta (se ha alcanzado la velocidad máxima que
corresponde al fenómeno de saturación). Es un transporte mediado.
Esta
difusión se da en el transporte de azúcares en la membrana plasmática de
hepatocitos.
b)
Transporte activo
Es
un transporte mediado que -a diferencia de la difusión facilitada- actúa en
contra de un gradiente de concentración y por eso requiere gasto de energía. Es
importantísimo en toda célula viva, ya que mientras los pasivos tienden a
homogeneizar el medio externo y el interno de la célula, los activos crean las
condiciones necesarias para que las células conserven su individualidad.
Este
transporte generalmente aumenta su velocidad al doble por cada 10 1C
de incremento de la temperatura, y puede ser inhibido por venenos (p. ej. el cianuro en la cadena respiratoria, o el malonato en el ciclo de Krebs).
Debe
haber acoplado al mecanismo de transporte un aporte energético. En muchas
células de mamíferos la energía se consigue por la hidrólisis del ATP. Esta
molécula tiene 2 uniones pirofosfóricas ricas en energía (12 kcal por unión).
ATP ==========> ADP + Pi +
12 kcal
Para
que esta hidrólisis suceda se requiere en el medio la presencia de una enzima
llamada ATPasa. En las membranas en las que se da
transporte activo existe esta enzima y en gran cantidad. Su actividad está
regulada por las concentraciones intra o extracelulares de las sustancias
transportadas activamente.
Un
ejemplo de transporte activo es el que realiza la bomba de Na+-K+,
que mantiene el potencial de membrana e interviene de forma muy activa en la
transmisión del impulso nervioso.