3) SISTEMAS DE MEMBRANA: membrana
plasmática (modelo del “mosaico fluido” de Singer y
Nicolson), retículo endoplasmático
liso y rugoso, complejo de Golgi, vacuolas y
lisosomas.
4. EL CITOSOL Y LOS RIBOSOMAS.
5. MITOCONDRIAS Y CLOROPLASTOS.
6. CENTRIOLOS, CILIOS Y FLAGELOS Y
MICROTÚBULOS.
7. EL NÚCLEO: membrana nuclear, nucleoplasma,
cromatina y nucleolos. Funciones del núcleo en reposo.
1. Introducción
Para estudiar la estructura de la célula eucariótica,
cuyas características y diferencias respecto a la célula procariótica señalamos
en el tema anterior, seguiremos el siguiente esquema:
1) La pared celular de las células vegetales.
2) Sistemas de membrana: membrana plasmática,
retículo endoplasmático liso y rugoso, complejo de Golgi, vacuolas y lisosomas.
3) El citosol y los ribosomas.
4) Mitocondrias y cloroplastos.
5) Centriolos, cilios y flagelos y
microtúbulos.
6) El núcleo.
2. La pared celular de las
células vegetales
Veíamos en el tema anterior que una de las diferencias
existentes entre la célula animal y la célula vegetal era la presencia en ésta
de una cubierta exterior rígida que denominamos pared celular. La pared celular
se sitúa por encima de la membrana plasmática. Por su rigidez actúa como sostén
mecánico en los tejidos vegetales.
En la constitución de las paredes celulares interviene
fundamentalmente la celulosa producida por la célula, pero las paredes
adyacentes se encuentran cementadas con pectina.
Las paredes celulares son muy complejas y están altamente
diferenciadas en algunos tejidos. Además, se disponen conforme a una secuencia
especial. Hablamos de paredes primarias, secundarias y terciarias, que se
depositan en capas superpuestas durante el crecimiento y la diferenciación
celular. La disposición de las microfibrillas (elementos constitutivos
de la mayoría de las paredes celulares) y la composición química diferencian
esos tres tipos de paredes celulares.
Las paredes primarias y secundarias están
compuestas principalmente por celulosa, a la que se pueden incorporar otros
materiales (lignina o suberina, cutina, ceras, etc.).
La pared terciaria se deposita, en algunos
tejidos, en el interior de la pared secundaria y posee una estructura especial
así como diferentes propiedades químicas y de tinción. Está compuesta
fundamentalmente por xilosa en lugar de celulosa.
La formación de las diversas paredes hay que considerarla
dentro del proceso de crecimiento y diferenciación de la célula. Durante el
crecimiento, las partes más externas y viejas de la pared celular están sujetas
a estiramientos intensos por los procesos de reorientación y aun de ruptura que
ocurren en las microfibrillas. Así se pueden producir los fenómenos de
desgarramiento, inclinación, crecimiento en mosaico, etc.
La pared celular determina en gran medida la forma de la
célula y se utiliza como un criterio para la clasificación de los tejidos
vegetales en parénquima, colénquima, fibras, etc.
La pared celular primaria está formada esencialmente por
microfibrillas de celulosa que pueden orientarse en todas las direcciones
dentro del plano de la pared, formando un retículo laxo que contiene grandes
cantidades de agua y componentes no celulósicos. Se admite que el crecimiento
de la pared primaria y la secundaria tiene lugar por aposición. Sin embargo, en
la pared secundaria las fibrillas son paralelas y se encuentran agrupadas más densamente.
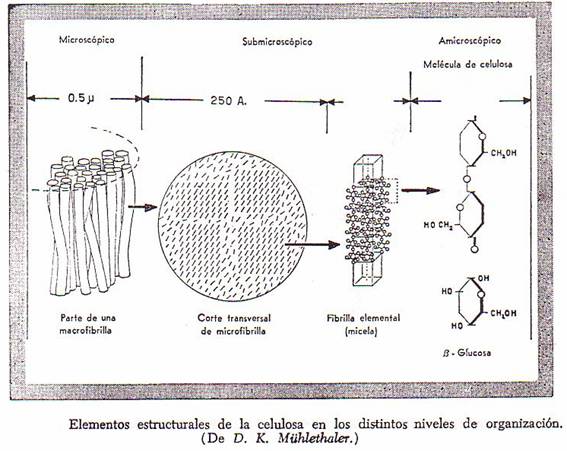
En la siguiente figura aparecen representados los
elementos estructurales de la celulosa en los distintos niveles de organización
dentro de la pared celular. Las macrofibrillas, visibles con el
microscopio óptico, están compuestas por microfibrillas de unos 250 Å de
diámetro, las cuales a su vez se encuentran constituidas por cerca de 2.000
cadenas de celulosa. Alrededor de 100 de estas cadenas están unidas entre sí en
fibrillas elementales o micelas, en las cuales existe una
organización molecular cristalina.
Queda decir por último que, si bien señalábamos al
principio la importancia de la pared celular como estructura ríguida para la
misma estructura de la planta, es también importante su papel estructural para
la célula considerada individualmente. No se trata sólo de conferirle la forma
característica, como ya hemos dicho: hay que entender que las células vegetales
están en contacto continuo con el agua -por la cual, a través de la raíz, toman
del suelo diversos nutrientes-, que presenta en disolución una concentración de
sales muy inferior a la que tiene la propia célula en su citoplasma. Esto,
unido al carácter semipermeable de la membrana plasmática, hace que las células
vegetales se encuentren sometidas a presiones osmóticas elevadas que, por
entrada de agua, llegarían a reventar la célula, si ésta no se encontrase
protegida por la "coraza" de la pared celular (en este caso la
"coraza" sería para impedir que se reventase la célula por la presión
que ejerce el agua desde dentro y no para defender a la célula de presiones
externas).
3) Sistemas de membrana:
membrana plasmática, retículo endoplásmico liso y rugoso, complejo de Golgi,
vacuolas y lisosomas.
a) Estructura y función de la membrana plasmática
La membrana plasmática constituye el límite de la célula,
a través del cual se realizan los intercambios con el medio extracelular.
Funciona como una barrera selectiva que incorpora determinadas sustancias del
medio e interviene en la eliminación de catabolitos o de sustancias elaboradas por
la célula.
No es una estructura estática, con función de
compartimentación, sino dinámica, cambiante, que además sirve de soporte de
enzimas.
Tiene unos 8 nm de espesor. Al micro electrónico con
muchos aumentos aparece como una estructura trilaminar con dos capas densas que
delimitan una capa central clara. En células animales la capa densa externa es
más gruesa que la interna.
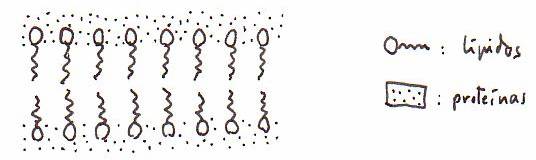
*Composición
química y estructura: la membrana plasmática está constituida por
lípidos y proteínas a las que se unen radicales glucídicos.
OVERTON (1895) observó que la membrana era atravesada
fácilmente por sustancias liposolubles. GORTER y GRENDEL (1925) idearon un
modelo en el que los lípidos formaban una bicapa. DAVSON y DANIELLI (1934)
precisaron más aún el modelo anterior: en la bicapa lipídica los grupos no
polares quedarían hacia dentro, limitados por los grupos polares de esos
lípidos, que quedarían hacia fuera (en contacto con el medio extracelular o con
el intracelular). Las proteínas constituirían una capa molecular a ambos lados
de la membrana, emparedando los lípidos.
Este modelo se consideró válido durante muchos años.
Actualmente se considera más probable que las proteínas
de la membrana sean globulares y se distribuyan de manera discontinua,
atravesando con frecuencia todo el espesor de la membrana. El modelo más
aceptado hoy es el del "mosaico fluido" de SINGER y NICOLSON
(1972):
Según este modelo, las proteínas constituyen unidades
separadas e independientes, y los fosfolípidos se ordenan en bicapas y forman
una matriz de cristales líquidos. En esta bicapa las moléculas de lípidos
pueden desplazarse lateralmente y así dotan a la bicapa de fluidez y
flexibilidad. Además confieren una resistencia eléctrica muy elevada a la
membrana y la hacen relativamente impermeable a las moléculas muy polares.
Las proteínas son globulares, según dice este modelo,
para interpretar su elevado contenido en á hélice. Algunas proteínas se encuentran
parcialmente empotradas en la fase lipídica y otras la atraviesan: esto dependerá
de la localización de los grupos R no polares de los aminoácidos.
De esta manera las proteínas formarían una estructura de
tipo mosaico en la fluida bicapa fosfolipídica. El mosaico no es fijo o
estático, ya que las proteínas pueden difundirse lateralmente.
Este modelo interpreta satisfactoriamente muchas
características y propiedades de las membranas biológicas: la diversidad en
contenidos de proteína por unidad de superficie membranosa; la asimetría de las
membranas naturales (por la distribución asimétrica de proteínas a ambos
lados); las propiedades eléctricas y de permeabilidad; y la elevada velocidad a
la que se mueven por la membrana algunas proteínas.
La membrana plasmática de las células eucarióticas posee
a menudo una cubierta exterior o glucocálix formada por cadenas
laterales oligosacáridas y por los oligosacáridos que constituyen los grupos de
cabeza de los glucolípidos de la propia membrana.
*Función:
además de limitante celular, la membrana tiene un papel importante en el
intercambio de sustancias entre la célula y el medio extracelular, y en la
recepción de información.
El transporte de agua e iones se realiza por medio de
canales hidrófilos cuya existencia se ha deducido indirectamente. En los axones
de las neuronas la membrana plasmática presenta 2 tipos de canales: una
"bomba" que transporta Na+ y K+ contra
gradiente de concentración consumiendo energía procedente del ATP; y otro tipo
de canales que permite el paso de iones a favor de gradiente de concentración y
se abre en respuesta a los cambios de voltaje a través de la membrana.
La membrana plasmática es además un receptor de señales
del medio extracelular. Presenta receptores específicos (probablemente
proteínas integrantes de la membrana) para la mayor parte de hormonas proteicas
y glicoproteicas que estimulan a las células.
*Diferenciaciones
de la superficie celular: en la superficie celular se encuentran con
frecuencia estructuras especiales relacionadas con las funciones que desempeñan
las células. Particularmente en las células epiteliales que revisten cavidades
(intestino, acinos y conductos glandulares):
-Microvellosidades: expansiones
citoplasmáticas que aumentan mucho la superficie celular libre. Facilitan la
absorción. Por ejemplo, en el epitelio intestinal.
-Pliegues basales: en la parte basal
-es decir, laopuesta a la luz- de
epitelios de revestimiento, con un importante papel en el transporte de iones.
Se trata de profundos pliegues de la
membrana, entre los que hay citoplasma con numerosas mitocondrias.
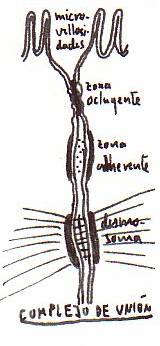
-Estructuras de unión entre células:
.Complejos
de unión (con una zona ocluyente o unión estrecha; una zona
adherente; y un desmosoma). Se dan en el epitelio cilíndrico del
intestino.
.Nexos
( o "gaps junction") en los que las membranas se aproximan
mucho pero no se fusionan, y en los que existen canales que unen los
citoplasmas de células adyacentes y por los que pueden pasar iones. Los nexos
se observan con frecuencia en fibras musculares (lisas y cardiacas).
b) Retículo endoplasmático rugoso
Descrito por PORTER y PALADE en 1952 como unas membranas
dispuestas en perfiles de dos membranas paralelas que entonces interpretaron
como túbulos. Un año después PALADE descubrió los ribosomas.
El RER está formado por cisternas con ribosomas en su
cara citoplasmática. Las membranas, lipoproteicas, forman bolsas
aplanadas (cisternas) y túbulos, con frecuencia interconectados.
En células con intensa síntesis proteica (células plasmáticas, acinares del
páncreas, etc.), el RER se dispone en forma de sacos aplanados y muy próximos,
por lo que en los cortes que se estudian al microscopio electrónico se ven como
"perfiles" de membranas paralelas.
El contenido de las cisternas depende de los
polipéptidos sintetizados.
Los ribosomas, adosados a la cara externa
(citoplasmática) se hallan dispuestos en polirribosomas. Se unen a la membrana
por la subunidad mayor.
Función. En cuanto a la función del RER, ésta es
la síntesis de proteínas que han de ser segregadas: al exterior (como las
células plasmáticas que fabrican las inmunoglobulinas), o al interior (como
ciertas enzimas líticas que son rodeadas por una membrana y constituyen los
lisosomas).
Los polipéptidos son sintetizados en la cara
citoplasmática (donde están los ribosomas) y deben pasar al interior de las
cisternas. Es probable que este paso se dé a través de ventanas que se abren
temporalmente en la membrana.
Para comprender cómo se completa la síntesis de proteínas
hay que tener en cuenta el Complejo de Golgi.
c) Retículo endoplasmático liso
Muy desarrollado en algunos tipos celulares: células
hepáticas, musculares, secretoras de hormonas esteroideas, secretoras de Cl-.
Presenta forma de red de túbulos interconectados y vesículas. Sin ribosomas
en la superficie citoplasmática.
En ocasiones RER y REL presentan conexiones. Pero ambos
son funcionalmente diferentes. Además de por no tener ribosomas, el REL se
diferencia del RER por estar constituido principalmente por túbulos, que
al ser cortados muestran forma de vesículas redondeadas, y no la forma de
perfiles alargados propia de las cisternas del RER.
Químicamente el REL está formado por proteínas y lípidos
que constituyen las membranas, y por proteínas enzimáticas.
Funciones. Son variables, según los tipos
celulares.
-En el hígado: guarda relación con el metabolismo
del glucógeno. También tiene una función desintoxicadora de drogas y
sustancias tóxicas (fármacos, carcinógenos, pesticidas) que son degradadas o
convertidas en no tóxicas por el REL de los hepatocitos.
-También se desarrolla más el REL en las células
que sintetizan esteroles (por ejemplo, las de la corteza suprarrenal).
-En el intestino: el REL de las
células absorbentes de la mucosa es donde se sintetizan los triglicéridos a
partir de los productos de digestión de las grasas.
-En el músculo estriado: el REL
almacena Ca2+ y lo libera para que se produzca la contracción
muscular.
d) Complejo de Golgi
Descubierto por CAMILO GOLGI (neurólogo italiano) en
1899, sirviéndose de la técnica de impregnación con una sal de plata con la que
realizaba sus estudios en el tejido nervioso.
*Localización.
En las neuronas, donde fue descubierto, se dispone preferentemente en torno al
núcleo. En otros tipos celulares tiene una clara polarización (en el epitelio
intestinal, en células musculares). En células plasmáticas se dispone en una
zona central próxima al núcleo.
Con frecuencia el complejo de Golgi rodea la parte de
citoplasma que contiene el centriolo y que se denomina citocentro.
*Ultraestructura.
Al micro electrónico se aprecian: cisternas aplanadas, vesículas y vacuolas.
En vertebrados las cisternas se apilan en número
variable. Cada cisterna suele adoptar forma de plato: con una cara cóncava, y
otra convexa y con el borde muy engrosado.
Rodeando el conjunto de cisternas existen vesículas
redondeadas con un material flocular que se condensa hasta originar los
gránulos de secreción.
El complejo de Golgi "crece" por su cara
convexa, que recibe vesículas del RER con material elaborado en el retículo. Y
esa membrana incorporada va desplazándose poco a poco a la cara cóncava de la
que se desprenden vacuolas y gránulos de secreción. Las membranas
de la cara convexa son algo más finas que las de la cara cóncava (5 y 7'5 nm
respectivamente).
En invertebrados y en células vegetales el complejo de
Golgi se dispone en "unidades" separadas, generalmente muy numerosas,
diseminadas por todo el citoplasma y formadas por cisternas y vesículas. Cada
unidad se llama dictiosoma.
*Función.
El complejo de Golgi juega un papel esencial en el proceso de secreción. Recibe
el producto sintetizado en el RER, completa la síntesis de las moléculas
añadiendo restos glicídicos, y concentra el material y constituye así los
gránulos de secreción que almacena o segrega al exterior.
Secreción de proteínas. Por ejemplo en células
acinares del páncreas: se sintetizan enzimas digestivas en el RER, de donde
pasan al complejo de Golgi mediante vesículas de superficie lisa que se generan
en los perfiles del RER. Se fusionan con las cisternas del Golgi y vierten su
contenido. De las cisternas del Golgi salen vacuolas con el material para
segregar, concentrado por un proceso de sustracción de agua a su paso por el
complejo de Golgi. Las vacuolas segregan su material fuera de la célula.
Secreción de glicoproteínas. Por ejemplo las
células plasmáticas: segregan inmunoglobulinas (anticuerpos). En este tipo de
secreción, la fracción proteica es sintetizada por el RER y de aquí es
transferida al Golgi, donde se completa la síntesis por adición de restos
glicídicos.
En células que segregan mucopolisacáridos sulfatados (p.
ej. las caliciformes del intestino) el complejo de Golgi, además de la síntesis
glicídica, realiza la incorporación del Ca2+.
En vegetales desempeña también la función de secreción de
la pared celular.
e) Vacuolas
Son los espacios o cavidades dedicados en la célula a
almacenar sustancias de reserva o de desecho. Mucho más desarrolladas en
vegetales que en animales (aunque en células vegetales jóvenes son muy
pequeñas).
En células vegetales. Almacenan sustancias de
reserva (almidón, grasas, proteínas, etc.) o pigmentos como los que
dan color a las flores. En muchos casos acumulan productos de desecho de
su propio metabolismo, muchos de los cuales son venenos más o menos activos:
resinas, cristales de oxalato cálcico, etc.
En células animales. Cuando las hay, pueden ser de
dos clases: vacuolas digestivas, dedicadas a la función digestiva; y vacuolas
pulsátiles, que regulan la cantidad de agua de la célula.
Además, en células animales se encuentran también
diversas inclusiones de sustancias de reserva (glucógeno, grasas, etc.).
f) Lisosomas
Aislados por DE DUVE (1949). Por centrifugación se
calculó que debían tener un diámetro de 0'2 a 0'8 ìm. Tenían un contenido elevado en fosfatasa
ácida y otras enzimas hidrolíticas. Por sus propiedades enzimáticas se les
denominó lisosomas.
Tienen una función digestiva (digestión intracelular). En
ellos se han reconocido más de una docena de enzimas hidrolíticas que, junto
con la fosfatasa ácida, tienen la propiedad de fragmentar compuestos biológicos
en un medio ligeramente ácido. Son característicos de células animales.
Tienen su origen en el complejo de Golgi, del que se
desprenden como vesículas cargadas de enzimas que se forman por gemación de los
sáculos de dicho orgánulo.
La membrana de los lisosomas no es atacada por las
enzimas que encierra, debido a que se halla recubierta interiormente por una
gruesa capa de glucoproteínas que impiden su autodigestión. Pero cuando las
células se ven privadas bruscamente de oxígeno, se produce la rotura de los
lisosomas con liberación de las enzimas y la consiguiente muerte de la célula
por autodigestión. Por el contrario, ciertas sustancias hormonales, como la
cortisona, estabilizan la membrana del lisosoma.
Se habla de lisosomas primarios cuando éstos se
encuentran libres en la célula sin actuar.
Los lisosomas secundarios son fruto de la fusión
de lisosomas primarios con las vacuolas de fagocitosis o las vesículas
pinocíticas que contiene materiales ingeridos. Por acción de las enzimas
hidrolíticas el material englobado (en macrófagos, por numerosos lisosomas
primarios) es digerido progresivamente.
4. El citosol y los
ribosomas
a) El citosol
El espacio existente entre los orgánulos citoplasmáticos
está lleno de un medio aparentemente amorfo que recibe el nombre de citoplasma
fundamental, citosol o matriz citoplasmática. Debido a su
aspecto hialino al microscopio óptico también se denomina hialoplasma.
Es soluble y experimenta cambios entre una fase viscosa y otra fluida,
probablemente debido a la distinta polimerización de las moléculas que lo
componen. En las células en movimiento predomina la fase líquida. En las
células fijas, la fase viscosa, aunque existen vías más fluidas por las que
circulan sustancias.
En la matriz citoplasmática se hallan las enzimas que
intervienen en el metabolismo de la glucosa. Además de proteínas, la matriz
contiene agua, iones, aminoácidos, ARN-t y la mayor parte de los metabolitos
resultantes del metabolismo.
Aquí se encuentran los orgánulos celulares y otros
componentes como el glucógeno, gotitas de lípidos, microfilamentos y
microtúbulos.
El glucógeno y las gotitas de lípidos
aparecen como sustancias de depósito, para el uso de la célula o para ceder a
otras. Los gránulos de glucógeno en el músculo son alargados y pequeños (15-30
nm), mientras que en el hígado presentan un tamaño mucho mayor (200-250 nm) y
forma redondeada. Los lípidos en el hígado y en otros órganos se presentan en
forma de pequeñas gotitas, mientras que en el tejido adiposo forman una gota
gruesa que ocupa casi toda la célula y desplaza el núcleo hacia la periferia.
Los microfilamentos son de naturaleza proteica.
Algunos formados por actina y miosina tienen función contráctil y son
particularmente abundantes en el músculo estriado (aunque también aparecen en
otros tipos celulares e intervienen en movimientos intracelulares).
b) Los ribosomas
Descubiertos por PALADE (1953). Son pequeñas partículas
con un diámetro comprendido entre 15 y 25 nm. Contienen ARN-r y proteínas en
proporción diferente (vid. tema 6) según sean de procariota o de eucariota.
Presentan dos subunidades que permanecen unidas en presencia de Mg2+.
El ARN-r se sintetiza esencialmente en el nucleolo.
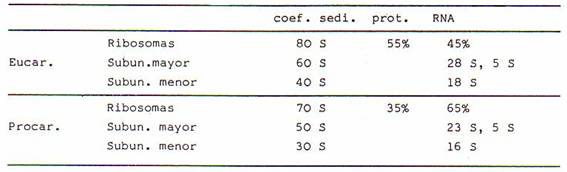
Generalmente se establece el tamaño de los ribosomas y de
sus subunidades en función de la velocidad a la que sedimentan en el tubo de la
centrífuga. Esta velocidad se mide en unidades S (de Svedberg). Los ribosomas
de células eucarióticas sedimentan a 80 S, y en ausencia de Mg2+ se
desdoblan en dos subunidades de 60 S y 40 S. Los ribosomas de procariotas,
mitocondrias y cloroplastos sedimentan a 70 S y están constituidos por dos
subunidades de 50 S y 30 S.
Los ribosomas de eucariotas contienen un 45% de ARN-r y
un 55% de proteínas. El ARN-r contenido en la subunidad mayor (de 60 S) tiene
una constante de sedimentación de 28 S, mientras que el ARN-r de la subunidad
menor (40 S) tiene una constante de sedimentación de 18 S. Los ribosomas de
procariotas tienen un 65% de ARN-r , con una constante de sedimentación de 23 S
y 16 S en la subunidad mayor y menor respectivamente. En la subunidad mayor
existe además un ARN-rde 5 S que no
guarda relación con el nucleolo.
La función de los ribosomas es la de sintetizar proteínas
(vid. tema 6). Durante esta síntesis, en los ribosomas se lee el código
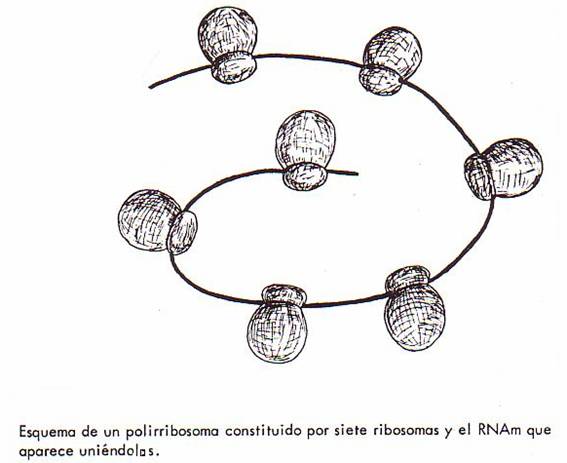
genético contenido en el ARN-m. Varios ribosomas aparecen entonces unidos por
una molécula de ARN-m formando los polirribosomas. El número de
ribosomas en un polirribosoma depende de la longitud del ARN-m que se esté
traduciendo:
en el caso de los polipéptidos
de la hemoglobina (con 150 aminoácidos por polipéptido) los polirribosomas
están integrados por 5 ribosomas.
5. Mitocondrias y
cloroplastos
a) Mitocondrias
Son orgánulos presentes en todas las células tanto
animales como vegetales, y cuya función es proporcionar a la célula la mayor
parte de la energía para sus funciones vitales, en forma de ATP. Estos
orgánulos pueden ser esféricos, ligeramente alargados o muy alargados con
aspecto filamentoso.
Su tamaño oscila entre 1 y 3mm. Esto hace que puedan ser observadas con el
microscopio óptico, aunque el estudio de su estructura detallada exija la
utilización del microscopio electrónico.
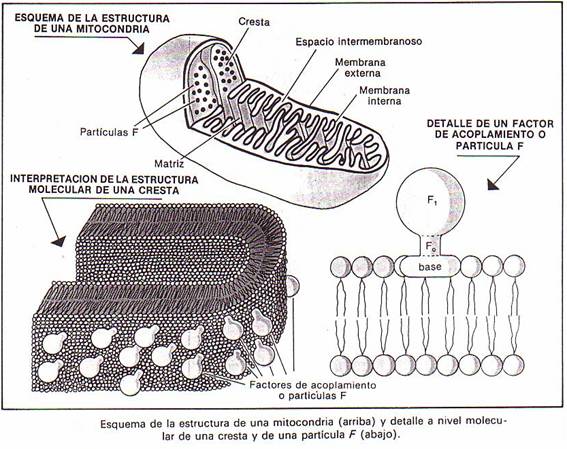
Son orgánulos huecos que constan de dos membranas con 60
Å de espesor cada una, separadas por un pequeño espacio llamado espacio
intermembranoso. La membrana externa es lisa. La membrana interna
forma una serie de invaginaciones o repliegues perpendiculares al eje
longitudinal de la mitocondria que reciben el nombre de crestas
mitocondriales. En las crestas se encuentran las enzimas necesarias para la
respiración celular.
El espacio que queda envuelto por la membrana interna se
denomina matriz mitocondrial y se encuentra ocupado por un líquido con
abundantes sustancias: enzimas que intervienen en el ciclo de Krebs, diversas
proteínas, y ácidos nucleicos (ADN y ARN). En ocasiones se observan densos
granos formados por lipoproteínas y ribosomas (mitorribosomas). Al
contener ADN, ARN y ribosomas, las mitocondrias sintetizan sus propias
proteínas: se trata de proteínas destinadas a la formación de la membrana
interna.
Al micro electrónico -y mediante la técnica de tinción de
contraste negativo- se ha visto que la membrana interna ,y por lo tanto las
crestas mitocondriales, se halla recubierta por unas partículas esféricas de
unos 90 Å de diámetro, unidas a la membrana por un pequeño pedúnculo de 35 Å de
diámetro y 50 Å de longitud. Estas partículas reciben el nombre de factores
de acoplamiento o partículas F y en cada mitocondria hay entre 104
y 105.
En cuanto a su estructura, las membranas responden al
tipo general de las membranas biológicas; pero carecen de colesterol y las
proteínas que se incrustan en la bicapa lipídica son más abundantes en la
membrana interna (constituyen el 80% de la membrana, mientras que los lípidos
representan sólo el 20%). Entre estas proteínas destacan las enzimas de
transporte electrónico de la cadena respiratoria.
En los factores de acoplamiento se pueden
distinguir tres partes: la F1 , de forma esférica;la F0, en forma de pequeño
pedúnculo cilíndrico (ambas -F1 y F0- sobresalen de la
membrana); y una base lipófila integrada en la propia membrana.
En cuanto a su función, en las mitocondrias tienen
lugar todas las reacciones del proceso de respiración celular. El proceso de
degradación de glucosa, como veremos, consta de tres fases: la glucolisis, el
ciclo de Krebs y la cadena de transporte electrónico asociada a la
fosforilación oxidativa. Pues en la matriz de las mitocondrias se encuentran
las enzimas que intervienen en el ciclo de Krebs, mientras que en las
crestas se encuentran las que intervienen en el transporte elctrónico y
en la fosforilación oxidativa (en esta intervienen, como veremos, las
partículas F). Precisamente para aumentar la superficie en la que se
desarrollan las acciones enzimáticas de la tercera fase del proceso, es por lo
que la membrana interna de las mitocondrias se encuentra replegada en forma de
crestas.
Lógicamente, en las células de organismos con respiración
anaerobia las mitocondrias son escasas o inexistentes.
El ADN mitocondrial está formado por una doble
hélice que en la mayor parte de los organismos estudiados forma una molécula
circular. La presencia de este ADN permite que las mitocondrias se reproduzcan
por bipartición.
Por lo que se refiere al ARN, existen los tres
tipos: ARN-r, ARN-m y ARN-t. Esto confiere cierta autonomía a las mitocondrias.
Los ribosomas se parecen (en sus coeficientes de
sedimentación y en su constitución) a los de procariotas, como ya hemos
estudiado.
b) Cloroplastos
Una de las características de las células vegetales es la
presencia de unos orgánulos específicos denominados plastos. De ellos los más
importantes son los cloroplastos, que contienen clorofila y son el asiento de
la fotosíntesis. En las plantas superiores los cloroplastos son de forma
ovalada, y su tamaño oscila entre 5 y 10 ìm de diámetro (son más grandes que las
mitocondrias). Su número varía bastante, pero en la mayoría de las células se
pueden contar entre 50 y 100.
Debido al color verde de la clorofila los cloroplastos se
pueden apreciar al microscopio óptico sin necesidad de tinción. Se perciben en
su interior unos granos cuya coloración es más intensa, debido a un acúmulo
mayor de clorofila. Fueron denominados grana.
En los cloroplastos de muchas algas se observan gránulos
esferoidales incoloros, denominados por SCHMITZ (1882) pirenoides. Son
acúmulos de almidón susceptibles de crecer.
Ultraestructura de los cloroplastos. Al micro
electrónico se observa que los cloroplastos están delimitados por dos membranas
(interna y externa), separadas por el espacio periplástico.
La membrana interna rodea la matriz o estroma,
que contiene un sistema de laminillas membranosas paralelas llamadas lamelas
o tilacoides.
Las lamelas recorren longitudinalmente el estroma
(recordemos que las crestas mitocondriales presentaban una disposición
transversal) y presentan expansiones en forma de monedas apiladas, de forma que
cada apilamiento constituye un granum (corresponde a uno de esos granos
más intensamente verdes que apreciábamos con el micro óptico). Actualmente se
denomina a la porción lamelar de los grana lamelas o tilacoides de los grana,
y a la porción rodeada de estroma lamelas o tilacoides estromales (éstas
pueden presentar perforaciones). Pero, como veremos, la composición de ambas es
diferente.
Las lamelas se originan, según la mayoría de los autores,
como invaginaciones de la membrana interna, pero en los cloroplastos bien
diferenciados las conexiones con ella se han perdido y sólo ocasionalmente se
aprecian. En el sistema lamelar se encuentran los pigmentos y enzimas que
intervienen en la fase luminosa de la fotosíntesis.
El desarrollo de tilacoides se ve favorecido por la
intensidad luminosa.
El sistema lamelar está presente en prácticamente todos
los cloroplastos. Sin embargo las algas tienen cloroplastos agranales. Y en
plantas superiores hay un conjunto de plantas denominadas C4 (que
fijan más CO2 que las demás) que contienen 2 tipos de cloroplastos:
las células del mesófilo tienen cloroplastos con grana, y las del parénquima
perivascular agranales.
En las membranas tilacoides sobresalen unas partículas
F semejantes en su estructura a las que hemos estudiado en mitocondrias
El estroma está ocupado por un líquido semejante
al del citoplasma y en el que se encuentran granos de almidón, macrogotas
lipídicas, una cierta cantidad de enzimas -las que intervienen en la fase
oscura de la fotosíntesis, que tiene lugar precisamente en el estroma- y
ADN, ARN y ribosomas (lo que hace que, igual que las mitocondrias, los
cloroplastos presenten cierta autonomía y sean capaces de multiplicarse por
bipartición).
La función de los cloroplastos es llevar a cabo la
función clorofílica o fotosíntesis. Ésta, como estudiaremos más adelante,
consta de 2 fases: fase luminosa (en la que intervienen pigmentos y enzimas
situados en el sistema lamelar) y fase oscura (en la que intervienen enzimas
contenidas en el estroma).
6. Centriolos, cilios y
flagelos y microtúbulos
a) El centriolo
El centriolo es un orgánulo presente en todas las células
animales. Generalmente, al micro óptico se aprecian dos gránulos (centriolos)
que constituyen el llamado diplosoma. En el diplosoma los centriolos se
disponen perpendicularmente. ALrededor del diplosoma se distingue una zona
esférica clara denominada centrosfera. De esta zona irradian un conjunto
de filamentos que, por su posición recuerdan a los rayos de un astro y reciben
el nombre de áster. Estos tres elementos (diplosoma, centrosfera y
áster) muy frecuentemente se sitúan en el centro de la célula y constituyen el citocentro
o centrosoma.
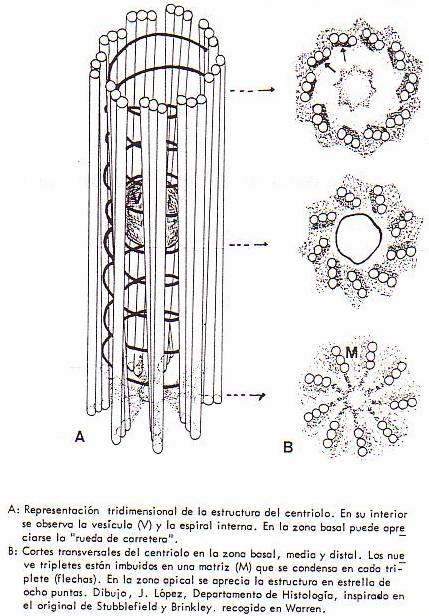
Con el microscopio electrónico se ha podido observar que
cada centriolo es un cilindro hueco (con dimensiones de 0'5-0'7 µm de longitud
por 0'25 µm
de diámetro) cuya pared está constituida por nueve conjuntos idénticos de tres
tubitos denominados microtúbulos, dispuestos tal y como representa la
figura. También los filamentos radiales del áster están formados por
microtúbulos.
En el interior del centriolo aparece una vesícula de 600
Å de diámetro cuya función y contenido nos resultan todavía desconocidos.
Por el interior del cilindro existe un filamento espiral
que recorre el centriolo en toda su longitud. Podría tratarse de ADN o ARN,
pero todavía no está claro.
Funciones. Una de las funciones del centriolo es
inducir la formación del huso acromático. Esta función es clara en las
células animales. En las vegetales -muchas de las cuales carecen de centriolos
visibles- también se forma el huso acromático.
La segunda función está relacionada con los cilios y
flagelos: inducen la formación de cilios y flagelos.
b) Cilios y flagelos
Aparecen en muchos seres unicelulares como órganos
locomotores. También aparecen en algunas células fijas de organismos pluricelulares.
La diferencia entre cilios y flagelos obedece tan sólo a
su número y tamaño, ya que desde el punto de vista estructural son semejantes.
Los cilios son más cortos y en general más abundantes; los flagelos, más largos
y escasos.
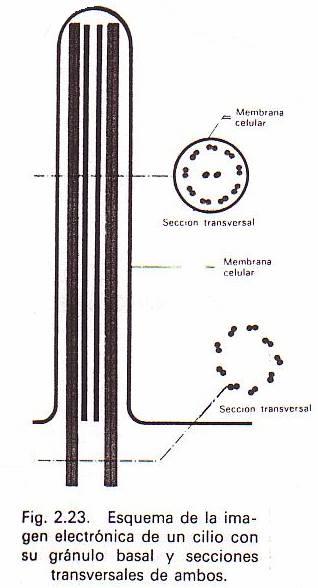
Cada cilio o flagelo tiene en su base una pequeña
granulación llamada gránulo basal o corpúsculo basal, de
estructura similar a la de un centriolo. El corpúsculo basal de los flagelos
suele ser más largo que el de los cilios. El flagelo o el cilio se encuentra
formado en su estructura externa por una prolongación de la membrana
plasmática. Tiene nueve pares de microtúbulos, continuación de los nueve
grupos que forman el corpúsculo basal. Pero tiene además otros dos microtúbulos
centralesque no tienen su equivalente en dicho corpúsculo.
Una vez más, los centriolos (en este caso constituyendo
el corpúsculo basal) desempeñan una actividad motora: rigen los movimientos de
cilios y flagelos (ya vimos que también dirigían los movimientos de los
cromosomas durante la división celular).
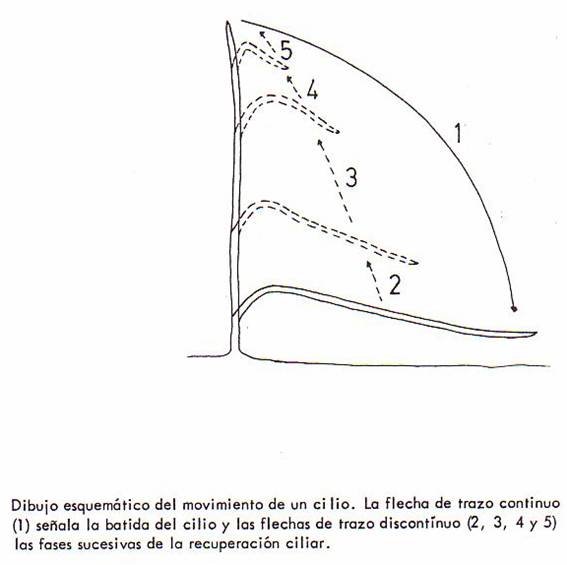
El movimiento de los cilios y flagelos es también
diferente. El movimiento de los cilios consta de dos etapas sucesivas.
Primero experimentan un golpe brusco y rápido (batida del cilio o fase
activa). A continuación ocurre la fase de recuperación, que es
lenta, con la cual se alcanza la posición original.
El movimiento de los flagelos es ondulante. En el
caso de los espermatozoides, ese movimiento, tridimensionalmente, describe un
cono. Igual que el de los cilios, el movimiento de los flagelos requiere ATP.
c) Microtúbulos
Los microtúbulos tanto de los centriolos como de los
orgánulos vibrátiles (cilios y flagelos) están constituidos por una proteína
llamada tubulina, cuyas moléculas se agrupan helicoidalmente por
polimerización y dan lugar a estas estructuras. Al ser polímeros de tubulina,
los microtúbulos son lábiles y pueden desintegrarse y reconstruirse con
facilidad a partir de las moléculas de tubulina que hay en el citoplasma. Esto
explica por qué cuando una célula ciliada o flagelada se divide, regenera con
facilidad los cilios y flagelos que les faltan a cada una de las células hijas,
y también explica la duplicación del centriolo y la formación y desaparición
del huso acromático en la división celular.
7. El núcleo
Es un corpúsculo bien definido que se halla inmerso en el
citoplasma y destaca con claridad en todas las células. Generalmente ocupa una
posición central, aunque en ocasiones se encuentra desplazado hacia la
periferia (como ocurre en las células vegetales por la presencia de grandes
vacuolas). Su forma suele ser esférica (no obstante aparecen núcleos
arriñonados, elipsoidales, en bastón, lobolados, etc.). En cada tipo de células
se mantiene constante la relación entre el volumen del núcleo y el del
citoplasma: es la relación nucleocitoplasmática.
El número de núcleos suele ser uno, pero no faltan casos
de células plurinucleadas que según su origen clasificamos como:
a) Sincitio: célula plurinucleada formada por
fusión de varias células uninucleadas.
b) Plasmodio: célula plurinucleada formada por una
repetida división nuclear que no ha sido acompañada de una división
citoplasmática.
En el núcleo interfásico -cuando la célula no se
encuentra en un proceso de división- distinguimos los siguientes componentes:
-membrana nuclear, que lo separa del
citoplasma;
-jugo nuclear o nucleoplasma;
-cromatina;
-nucleolos.
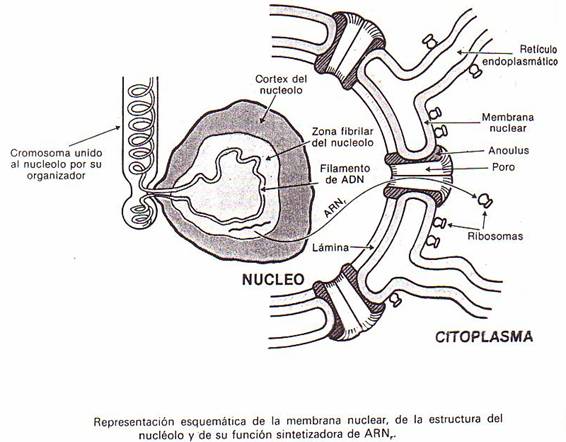
a) Membrana nuclear
Es una doble membrana formada por dos finísimas
hojasentre las que queda un espacio que se continúa con el del retículo
endoplasmático (dicha membrana no es más que una prolongación del retículo
endoplasmático que rodea el núcleo).
No forma una cubierta continua: en varios puntos de la
membrana existen interrupciones por la presencia de unos poros que
comunican el núcleo con el citoplasma. Bordeando cada poro de la membrana
nuclear se encuentran ocho subunidades o piezas dispuestas en anillo que
reciben el nombre de annulus y que, lógicamente, disminuyen el tamaño
del poro.
Sobre la membrana nuclear interna se ha descubierto
también una capa de material denominada lámina.
b) Nucleoplasma
Es la fase más o menos líquida, incoloreable, con
proteínas y otras sustancias disueltas o en suspensión. En su seno se
encuentran inmersos la cromatina y el o los nucleolos.
c) Cromatina
Es el componente más abundante de los núcleos. Se tiñe
con los colorantes básicos. Las masas de cromatina muy teñidas (heterocromatina)
aparecen al microscopio electrónico como zona muy densas a los electrones. Las regiones de cromatina que se tiñen
menos (eucromatina) son poco densas a los electrones.
La cromatina es el constituyente básico de los cromosomas
eucarióticos tanto en la interfase como durante la mitosis. Según parece, las fibrillas
de cromatinaestán constituidas por
la asociación entre moléculas lineales de ADN y proteínas
nucleares, fundamentalmente histonas ; también pueden existir pequeñas
cantidades de ARN. Así, la cromatina estaría formada por nucleoproteínas, cuyo
grupo prostético sería el ADN. Las proteínas del grupo proteico pueden ser
básicas o no básicas (ácidas). Las proteínas básicas son fundamentalmente las histonas
(con bajo peso molecular y una elevada proporción de aminoácidos básicos, tales
como la argininay la lisina).
Las histonas se unen al ADN por el ácido fosfórico, ya que mientras éste
presenta cargas negativas, los aminoácidos de las histonas están cargados positivamente a pH fisiológico. Las proteínas
no básicas parecen ser las responsables de que los cromosomas mantengan su
estructura, y por eso se las denomina proteínas estructurales.
Al microscopio electrónico se han podido observar las
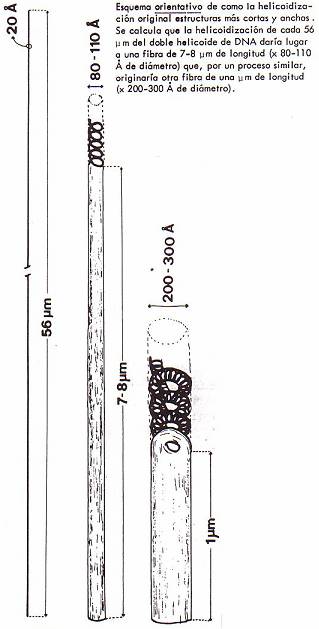
fibras de cromatina (microfibrillas). DU DRAW interpretó que cada fibrilla
consta de un sólo ADN de doble hélice (diámetro, 20 Å) asociado a
histonas. La cadena de ADN (que da continuidad a la fibra), sufriría un
proceso de helicoidización "enrollándose" sobre sí misma, y se
originarían así estructuras cada vez más cortas y más anchas, estabilizadas por
las proteínas asociadas al ADN.
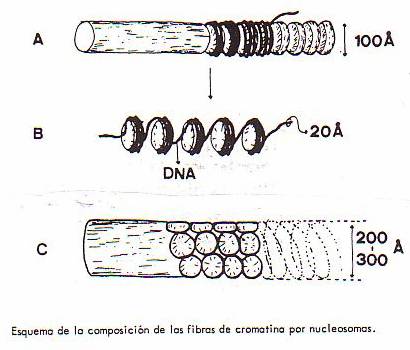
Los estudios posteriores han llevado a afirmar que la cromatina,
por lo menos en las zonas heterocromáticas, está formada por la asociación
longitudinal de pequeños cuerpos semejantes a una píldora, denominados nucleosomas
y cuyas dimensiones son: 10'5 x 5'5 nm. Cada nucleosoma consta de ocho
moléculas de histona asociadas a la cadena de ADN (el cual "rodearía"
a cada grupo de ocho histonas y conectaría cada nucleosoma con sus vecinos). Se
calcula que cada grupo de histonas se asocia a doscientos pares de nucleótidos
de la doble hélice de ADN.
El complejo ADN-nucleosomas da lugar a las distintas
fibras de cromatina (de 100 ó de 200-300 Å) que se pueden observar al
microscopio electrónico, según el modo de "empaquetarse" (en el nivel
de "empaquetamiento" correspondiente a las fibras de 200-300 Å, cada
vuelta del helicoide estaría formada por seis nucleosomas aproximadamente).
d) Nucleolos
El nucleolo es un corpúsculo más o menos esferoidal,
visible fácilmente debido a su tamaño (de 1 a 7mm) y a que su viscosidad y refringencia son
mayores que las del resto del núcleo. Es frecuente que exista más de un
nucleolo (dos o tres).
En el nucleolo se distinguen dos zonas: una de aspecto fibrilar
que ocupa la parte central, y otra de aspecto granular (también llamada córtex)
que rodea la anterior. Ambas zonas contienen ARN y proteínas (5-10% y 90%
respectivamente).
Los nucleolos están unidos a unas zonas específicas del
ADN cromosómico que se denominan organizadores nucleolares y que
contienen los genes que codifican para el ARN-r. Contienen múltiples copias de
estos genes: se trata de un ADN repetitivo. Los organizadores
nucleolares se sitúan generalmente en las zonas correspondientes a
constricciones secundarias dentro de los cromosomas.
En los nucleolos
tiene lugar la síntesis del ARN ribosómico.
Enumeración de las funciones del núcleo en reposo
1) Almacenamiento de la información
genética.
2) Transcripción de la información
hereditaria: síntesis de ARN-m, ARN-t y ARN-r, y transporte al
citoplasma.
3) Mecanismo de control que conduce a
la expresión de determinados genes y a la represión de
otros en cada célula particular en un momento concreto y que son de gran
importancia en el proceso de diferenciación y especialización celular. Aunque
los mecanismos de regulaciónde la expresión génica no están claros, las proteínas
no histonas parecen tener una gran importancia en el control de la
transcripción. También se piensa que otro factor importante en el control se da
a nivel post-transcripcional, mediante el transporte selectivo de los
ARN-m sintetizados al citoplasma.
4) Capacidad de responder a
"informaciones" de cambios intra o extracelulares, que le llegan
desde el citoplasma, mediante cambios en los mecanismos de control.
5) Replicación de la información
hereditaria antes de cada división celular.