2. LÍPIDOS. Características generales. Funciones. Clasificación.
Lípidos complejos o saponificables (ácidos grasos y saponificación): acilglicéridos, fosfoglicéridos,
esfingolípidos y ceras. Lípidos simples o insaponificables: terpenos, esteroides y prostaglandinas.
1.
Glúcidos
Los glúcidos
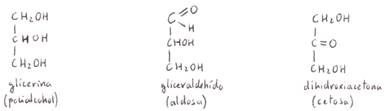
son principios inmediatos orgánicos formados por C, O y H en la proporción Cn(H2O)n,
por lo que antiguamente fueron llamados "hidratos de carbono" o
"carbohidratos". Resultan de sustituir en un polialcohol, mediante deshidrogenación,
uno de los grupos funcionales alcohol (-OH), por otro grupo funcional
aldehídico (-COH) o cetónico (-CO-).
Dentro de los glúcidos distinguiremos para su estudio tres grupos:
monosacáridos, disacáridos y polisacáridos. Por su sabor dulce, los dos
primeros se denominan "azúcares".
a) Monosacáridos
Son azúcares o glúcidos sencillos de fórmula general Cn(H2O)n,
siendo n, ordinariamente, 3, 4, 5, 6 ó 7 (rara vez más). No son hidrolizables (es decir, no se descomponen por hidrólisis
en azúcares más sencillos). Presentan isómeros ópticos (propiedad que
estudiaremos después). Los monosacáridos tienen sabor dulce (azúcares), color
blanco, son solubles en agua y tienen poder reductor debido al grupo carbonilo
(aldehído o cetona) que puede oxidarse para dar lugar a un grupo ácido (-COOH).
Según cuál sea el grupo
funcional (aldehído o cetona), distinguimos dos familias de monosacáridos: aldosas y cetosas.
Según cuál sea el número de C de
la molécula tendremos: triosas, tetrosas,
pentosas, hexosas y heptosas.
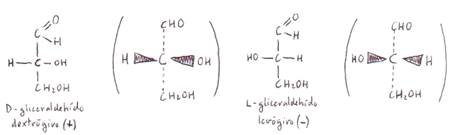
La aldosa
de 3 C (gliceraldehído) presenta un "C
asimétrico", es decir, con cada una de sus 4 valencias saturada por un
grupo químico diferente: el C2. Los compuestos con este tipo de
carbonos son ópticamente activos, es
decir, sus disoluciones desvían el plano de la luz polarizada. El sentido de la
desviación depende de la posición que ocupan los radicales -H y los -OH
respecto al C asimétrico al que se encuentran unidos: si en una posición
desvían el plano hacia la derecha (sustancia dextrógira), en la otra lo
desviarán hacia la izquierda (sustancia levógira). Se trata de la isomería
óptica.
Una mezcla equimolecular
de isómeros ópticos contrarios, será ópticamente
inactiva, es decir, no hará girar en ningún sentido el plano de la luz
polarizada.
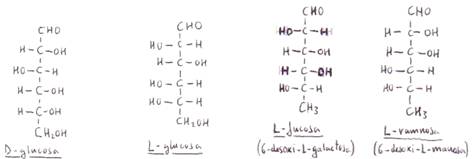
Los isómeros que presentan los glúcidos se dividen en dos series: serie D y serie L, según
cuál sea la posición del grupo -OH unido al C contiguo al grupo alcohólico
primario.
No todos los azúcares de la
serie D son dextrógiros, ni levógiros los de la serie L. Por eso la notación
completa de estas sustancias incluye el signo referente a la serie a la que
pertenecen y el correspondiente a su actividad óptica:
D (+) gliceraldehídoyL (-) gliceraldehído
La mayoría de los glúcidos naturales pertenecen a la serie D.
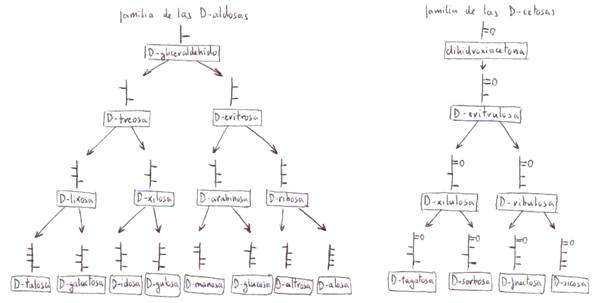
Las D-aldosas
de mayor importancia biológica son el D-gliceraldehído,
la D-ribosa, la D-glucosa, la D-manosa y la
D-galactosa.
Las cetosas
más importantes biológicamente son la dihidroxiacetona,
la D-ribulosa y la D-fructosa.
La forma habitual de la glucosa
hallada en la naturaleza es dextrorrotatoria o
dextrógira ([α]D = +52'71), mientras que la forma
corriente de la fructosa es levorrotatoria o levógira
([α]D = -92'41). Sin embargo, ambas pertenecen
a la serie D.
Las aldosas
y cetosas de la serie L son imágenes especulares de
sus correspondientes formas D: isomería espacial o estereoisomería.
En la naturaleza se encuentran
L-azúcares, pero no son tan abundantes como los D-azúcares. Entre los más
importantes se encuentran la L-fucosa y la L-ramnosa (desoxiazúcares) y la L-sorbosa.
Pero estas estructuras lineales
de monosacáridos sólo se dan en estado sólido (cristalino), pues se ha
comprobado que esos azúcares en disolución adoptan una estructura de cadena
cerrada o cíclica (con excepción de triosas o tetrosas, que mantienen su estructura lineal).
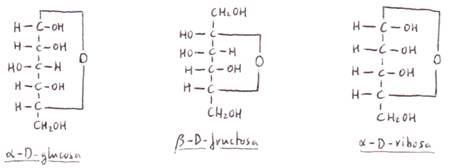
Las estructuras cíclicas se
producen por la unión del C que lleva el grupo aldehídico o cetónico
con un C que lleva un grupo alcohol dentro de la misma molécula. Se trata de un
enlace hemiacetal. En las hexosas
reacciona el grupo carbonilo (aldehído o cetona)con el
hidroxilo del C5, mientras que en las pentosas
lo hace con el hidroxilo del C4.
La formación del enlace hemiacetal constituye el C1 (o el C2
en las cetosas)en un nuevo C
asimétrico, lo que da lugar a los isómeros α y β (denominados anómeros) según cuál sea la posición del -OH unido a
ese C). Dicho C se denominará C anomérico.
Los anómeros
presentan una actividad óptica específica. Así la D (+) glucosa recientemente
disuelta tiene un poder rotatorio de +1091, que decrece hasta alcanzar un
valor de +52'51. Este fenómeno, que se conoce
como mutarrotación, se debe a la coexistencia
en el equilibrio de las formas α y β, cuyas rotaciones son
respectivamente +1091 y +19'81.
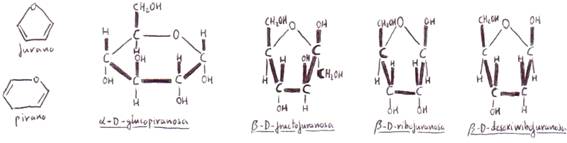
Las fórmulas cíclicas pueden ser
representadas de manera espacial mediante las fórmulas en perspectiva
propuestas por Haworth. De estas fórmulas, las que
presentan anillos pentagonales se denominan furanosas
(cetohexosas y aldopentosas),
y las que presentan anillos hexagonales (p. ej. las aldohexosas) reciben el nombre de piranosas.
Aunque en las formas cíclicas se
pierda la función aldehído o cetona, no desaparece el poder reductor, debido a
que en toda solución se da un equilibrio en el que coexisten formas cíclicas
con pequeñas cantidades no cíclicas.
*Funciones: en cuanto a las
funciones de los monosacáridos hay que destacar como principal la energética.
Es el caso de la glucosa, que entra en la ruta denominada glucolisis,
que se prolonga -en presencia de oxígeno- con el ciclo de Krebs
y la cadena de transporte de electrones. También la fructosa entra en dicha
ruta. Y otros azúcares (como la galactosa, etc. se transforman en glucosa para
seguir el mismo proceso. Se trata de una energía de aporte rápido, pues al ser
solubles en agua, los azúcares son llevados con facilidad a través de la sangre
a las diferentes partes del cuerpo. Pero los monosacáridos con función
energética se almacenan normalmente en forma de polisacáridos (que ya
estudiaremos).
Otros monosacáridos se asocian
entre sí para dar lugar a polisacáridos con función estructural (que
también estudiaremos). De igual forma, las pentosas
ribosa y desoxirribosa asociadas a grupos fosfato cumplen funciones de
"engarce" para las bases nitrogenadas en la estructura de los ácidos
nucleicos.
Por último, recordar que las pentosas también forman parte de coenzimas, como los
adenosín-fosfatos y los piridín-nucleótidos.
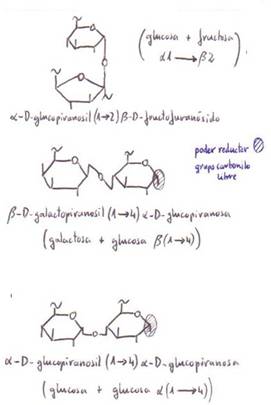
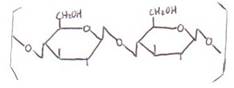
b) Disacáridos
Son también de sabor dulce,
cristalizables y solubles en agua, pero muchos de ellos no tienen poder
reductor, como ya veremos.
Se forman por unión de 2
moléculas de monosacárido mediante un enlace O-glicosídico
con desprendimiento de una molécula de agua.
El enlace O-glicosídico se establece al reaccionar un
grupo hidroxilo (-OH) de un monosacárido con otro grupo de idéntica naturaleza
de otra molécula. Se forma un puente de O y se desprende H2O:
monosac.-OH+HO-X====>monosac.-O-X+H2O
En el caso de los disacáridos,
ambos grupos -OH pertenecen a monosacáridos:
C6 H12O6+C6
H12O6=========>C12 H22O11+H2O
El enlace O-glicosídico
puede establecerse:
1) Entre los -OH de los grupos
carbonilo de ambos monosacáridos.
2) Entre el -OH del grupo
carbonilo de un monosacárido y otro -OH no del grupo carbonilo del segundo
monosacárido.
En el primer caso quedan
anulados ambos grupos carbonilos y por eso el disacárido resultante carece de
poder reductor.
En el segundo caso permanece el
poder reductor, al mantener uno de los monosacáridos su grupo carbonilo sin
bloquear.
(la
mezcla resultante se denomina azúcar invertido, por su actividad óptica)
*Lactosa:
-Azúcar de la leche.
-Hidrólisis ácida (intervienen β-galactosidasas,
en células del intestino). Los
niños intolerantes a la leche
presentan carencia total o parcial de galactosidasas.
-Sí es reductor.
*Maltosa:
-Azúcar de la malta.
-Producto intermediario del
catabolismo del almidón.
-Se rompe por la
α-glucosidasa.
-Sí es reductor.
c) Polisacáridos
Resultan de la unión de n
moléculas de monosacáridos por enlaces glicosídicos
con desprendimiento de n-1 moléculas de agua.
Los polisacáridos no
cristalizan. Tampoco forman verdaderas soluciones en el agua, sino coloides. Por hidrólisis se descomponen en disacáridos y,
después, en monosacáridos.
*Almidón: es la principal reserva
alimenticia de los vegetales. Muy abundante en tubérculos, semillas, etc.
Se almacena en las vacuolas de la célula vegetal. Con el agua forma una
solución coloidal denominada "engrudo de almidón", que se
colorea en azul al añadirle tintura de yodo. Presenta entre 1.000
y 5.000 unidades de glucosa.
El almidón es en realidad una
mezcla de 2 componentes: amilosa y amilopectina.
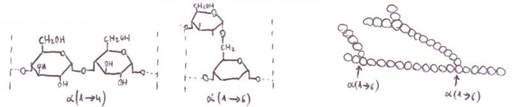
-Amilosa:
enlaces α(1==>4) entre las moléculas de
glucosa. Sin ramificaciones. Estructura helicoidal: cada vuelta con 8-10 restos
de glucosa. Con 200 moléculas de glucosa como mínimo. Soluble en agua caliente.
-Amilopectina:
enlaces α(1==>4) con ramificaciones
α(1==>6) cada 18-22 restos. Con unas 1.000 moléculas de glucosa.
Insoluble en agua caliente (de ahí el aspecto grumoso del almidón en agua).
El
iodo:en la amilasa produce un
color azul, ya que se integra en la estructura.
en la amilopectina, color pardo
(no se integra en la estructura).
-Enzimas amilolíticos
(hidrolasas del almidón): hidrolizan el almidón según
la secuencia:
almidón ===> dextrina ===>
maltosa ===> glucosa
La dextrina está constituida por
la parte de molécula respetada por las enzimas que no hidrolizan el enlace
1==>6: a partir de las ramificaciones.
Las amilasas hidrolizan
enlaces 1==>4 y dan lugar a restos de maltosa. Las enzimas desrramificantes hidrolizan los enlaces 1==>6. La maltasa
hidroliza la maltosa y da lugar a restos de glucosa.
*Glucógeno: es la reserva
alimenticia en animales. Por eso se le denomina también "almidón
animal". Se almacena en el hígado principalmente. También en músculo. En
el hígado se producen variaciones diarias de hasta un 20% debido a los procesos
de glucogenogénesis (síntesis de glucógeno) y glucogenolisis (degradación).
Está constituido el glucógeno
por restos de glucosa unidos por enlaces α(1==>4)
y ramificaciones α(1==>6) cada 10-12 restos. Está más ramificado que la
amilopectina: se empaqueta y almacena más fácilmente.
La frecuencia de las
ramificaciones es mayor en el interior de la molécula que en el exterior.
Puede alcanzar dimensiones de
5.000-30.000 restos de glucosa.
En la síntesis de glucógeno
intervienen la glucógeno sintetasa y la enzima
ramificante 1,4-α-glucano (para las
ramificaciones 1==>6).
En la degradación, la glucógeno
fosforilasa (para los enlaces 1==>4) y la enzima
desramificante (que deshace las ramificaciones
1==>6 y las recoge en forma lineal, 1==>4, para que actúe la glucógeno fosforilasa).
-)Por qué el glucógeno en lugar de
las grasas para almacenar energía?
1) Las grasas son más difíciles de movilizar
(para energía rápida no sirven).
2) Las grasas no se pueden utilizar como
fuente de energía en ausencia de O2.
3) Las grasas no son sustrato para la
obtención de glucosa, y hay células que requieren glucosa.
-)Por qué glucógeno en lugar de
glucosa libre?
1) Mientras que [glucógeno] es aprox.
0'01μM, con glucosa libre aumentaría la [glucosa] hasta 400mM y se
alteraría la Π (presión osmótica).
2) Al estar ramificado se almacena
mejor. Además la ramificación dificulta que se agote el glucógeno.
3) La actividad de la glucógeno sintetasa aumenta al disminuir el tamaño del cebador
(resto de glucógeno), por lo que se dificulta la hidrólisis total del glucógeno
celular. Y disminuye la actividad de la glucógeno sintetasa
al aumentar mucho las dimensiones del cebador (por lo que no se almacena
glucógeno en exceso).
Recién comido aumenta la glucogenogénesis del organismo. 2 ó 3 horas después,
aumenta la glucogenolisis. Pero el organismo humano regula
la glucemia entre 0'8 y 1'2 g de glucosa/l de sangre.
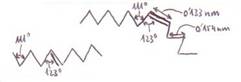
*Celulosa: tiene función
estructural en las células vegetales (forma parte de la pared celular). Sin
ramificaciones. Moléculas de β-D-glucosa (en lugar de la α-D-glucosa
de almidón y glucógeno) con enlaces 1==>4.
Entre 3.000 y 15.000 restos de
glucosa por cadena.
Además de los enlaces covalentes
β(1==>4) entre los restos de glucosa de una
misma cadena, también se establecen puentes de H y fuerzas de Van derWaals entre las cadenas, por
lo que la estructura se estabiliza más.
Los haces de cadenas constituyen
microfibrillas, que a su vez se agrupan en láminas,
y éstas se amontonan en capas (se presentan las microfibrillas
de una lámina cruzadas respecto a las de la lámina contigua).
La hidrólisis del enlace β(1==>4) entre moléculas de glucosa se realiza por
acción de las celulasas. Estas enzimas se
hallan presentes en algunas bacterias. Como algunas de esas bacterias se
encuentran presentes en el estómago de los rumiantes, estos animales pueden alimentarse
de celulosa ya que dichas bacterias colaboran en el proceso digestivo
descomponiendo la celulosa en restos de glucosa.
*Otros polisacáridos: sin ser
exhaustivos, conviene citar la función estructural de los polisacáridos que
forman parte de los péptidoglicanos en la
pared bacteriana; y de la quitina (polímero de unos 100 restos de N-acetilglucosamina) que recubre
el exoesqueleto de artrópodos.
2.
Lípidos
a) Características generales
*Son sustancias untuosas al tacto.
*Insolubles en H2O (aunque
algunos tienen una parte polar y otra apolar: son anfipáticos, y pueden actuar como detergentes:
dañinos para las sustancias biológicas).
*Solubles en los llamados
"disolventes orgánicos" como el cloroformo, éter, alcohol, sulfuro de
carbono, xilol, etc.
b) Funciones
Aunque iremos estudiándolas
conforme veamos cada clase de lípidos, podemos enumerar ahora las siguientes:
*Fuente de energía: es el caso
de los triglicéridos que se almacenan en el tejido adiposo. Por oxidación: -lípidos ===> 9'4 Kcal/g
-glúcidos
==> 4'1 Kcal/gdebido a que el C de los lípidos está más reducido que el de los glúcidos.
Además, su "almacen" (tejido adiposo) no es limitado como el del
glucógeno (hígado).
*Función estructural: en
membranas citoplasmáticas: ácidos grasos y colesterol.
*Aislante térmico: capas de
grasa de animales que viven en zonas frías.
*Función impermeabilizante: en
pelos, lana, plumas, hojas...
*Actividad biológica específica:
hormonas, vitaminas y prostaglandinas.
c) Clasificación
Los lípidos se han clasificado
de muy diversas maneras. Nosotros adoptaremos el criterio de clasificación
basado en la estructura de sus esqueletos.
*Lípidos complejos
(saponificables): contienen ácidos grasos como componentes. Se llaman también
lípidos saponificables porque producen jabones (sales de los ácidos
grasos) por hidrólisis alcalina. Difieren en la estructura del esqueleto al que
se hallan unidos -covalentemente- los ácidos grasos:
-acilglicéridos
==============> glicerina
-fosfoglicéridos
=============> 3-fosfato de glicérido
-esfingolípidos
==============> esfingosina
-ceras ====================>
alcoholes no polares de p.m. elevado
*Lípidos sencillos (insaponificables): no contienen ácidos grasos en su
estructura:
-terpenos
-esteroides
-prostaglandinas
d) Lípidos complejos
(saponificables)
Como hemos dicho, los lípidos
complejos o saponificables se caracterizan por presentar ácidos grasos en su
estructura.
*Ácidos grasos: son moléculas
consistentes en una cadena hidrocarbonada larga con
un grupo carboxilo terminal.
Estructuralmente las moléculas
de ácidos grasos adoptan la forma de zig-zag.
La cadena hidrocarbonada
puede ser saturada (toda con enlaces sencillos) o presentar insaturaciones
(uno o más dobles enlaces, rara vez triples enlaces). Así los ácidos grasos
difieren entre sí por la longitud de la cadena y por el número y posición de
los dobles enlaces.
A menudo se simbolizan con una
notación taquigráfica que indica: 11) el número de dobles enlaces y 21) el número, situación y
configuración de los dobles enlaces.
Se pueden hacer algunas
generalizaciones sobre los ácidos grasos presentes en las plantas
superiores y en los animales:
-Los más abundantes poseen un
número par de C.
-Las cadenas son de 14 a 22 C,
aunque predominan las de 16 y 18 C.
*Ácidos grasos saturados más corrientes:palmítico (C16)
esteárico (C18)
*Ácido graso insaturado más frecuente:oléico (C18:1
Δ9)
-Los ácidos grasos insaturados
predominan sobre los saturados.
-En los monoinsaturados
de los organismos superiores el doble enlace suele darse entre los C9
y C10.
-En los poliinsaturados,
un doble enlace suele darse entre C9 y C10 y los
siguientes se encuentran separados por un grupo metileno (-CH=CH-CH2-CH=CH-). Sólo en unos pocos tipos de ácidos grasos
vegetales se encuentran dobles enlaces conjugados (-CH=CH-CH=CH-).
-Los dobles enlaces de los
ácidos grasos insaturados que hay en la naturaleza suelen aparecer en
configuración geométrica cis.
-Muy pocos
la presentan trans.
En adelante, si no indicamos
otra cosa, se sobreentiende que los dobles enlaces son cis.
En bacterias los ácidos grasos
son menos y más simples: ácidos grasos saturados de C12 a C18,
y ácidos monoinsaturados C16 y C18.
No se han encontrado poliinsaturados.
-Algunos ácidos grasos
presentes en la naturaleza:
1) Son bipolares: cadena hidrocarbonada hidrófoba (o lipófila)
grupo -COOH terminal hidrófilo (o lipófobo)
Y de acuerdo con esas afinidades
se empaquetan en medio hidrófilo y en medio hidrófobo.
2) En disolución tienden a dimerizarse (proceso exergónico)
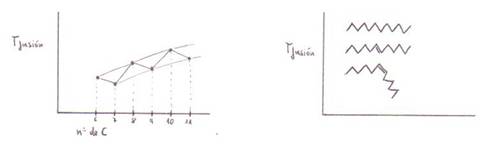
3) Los puntos de fusión son:
mayormenor
C paresC impares
saturadosinsaturados
insaturadostransinsaturados cis
4) Los puntos de ebullición son
elevados, y aparecen dímeros incluso en estado gaseoso. Aumentan los puntos de
fusión con el n1
de C. En cambio influye poco en esto el n1 de insaturaciones.
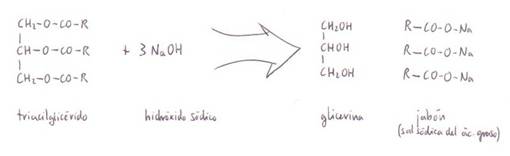
5) Saponificación. Se produce por
reacción del grupo -COOH terminal de los ácidos grasos con cationes
monovalentes de las bases (KOH ó NaOH), en caliente.
Se obtienen las sales sódicas o potásicas de los ácidos grasos, también
llamadas jabones. Son de interés industrial.
El jabón hace que las pequeñas
gotitas de grasa que se forman al agitar las grasas en el agua, sean más
estables: por su parte hidrófila se pone en contacto con el agua, y por su
parte hidrófoba se mezcla con las grasas. De forma similar actúa la bilis,
de forma que favorece la digestión de las grasas por acción de la lipasa
(la lipasa hidroliza las grasas pero no da lugar a jabón, sino a ácidos grasos
libres que pasan a la β-oxidación.
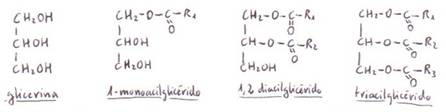
*Acilglicéridos
(o glicéridos): son ésteres de ácidos grasos con el alcohol glicerina.
Si los 3 grupos alcohol de la
glicerina se encuentran esterificados, la estructura
se llama triacilglicérido (es preferible no
utilizar el nombre de triglicérido porque químicamente es inexacto). Si los triacilglicéridos son sólidos a temperatura ambiente se les
denomina grasas, y si son líquidos, aceites (ver influencia de
las aturaciones e insaturaciones
y del n1
de C en el punto de fusión de los ácidos grasos).
Los diacilglicéridos
(o diglicéridos) y los monoacilglicéridos
(monoglicéridos) también se encuentran en la naturaleza,
pero en cantidades mucho menores. Aparecen sobre todo como pasos intermedios en
la oxidación de los triacilglicéridos.
Los triacilglicéridos
que contienen una sola clase de ácido graso en las 3 posiciones se denominan triacilglicéridossimples. Si contienen 2 ó 3 ácidos
grasos diferentes se llaman triacilglicéridosmixtos.
-Propiedades de los triacilglicéridos:
1) Su punto de fusión depende de los ácidos
grasos componentes (aumenta con el n1 y la longitud de los ácidos grasos componentes).
2) Son insolubles en agua. En cambio, los monoacilglicéridos y los diacilglicéridos,
por sus grupos hidroxilo libres, presentan cierta polaridad y forman micelas.
3) Hidrólisis:
-Al hervirlos con
ácidos o con bases. Hidrólisis con álcalis: saponificación (como
resultado se obtiene una mezcla de jabón y glicerina).
-Por acción de las
lipasas (jugo pancreático)
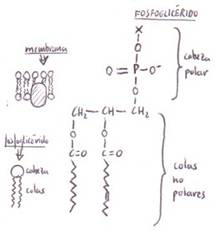
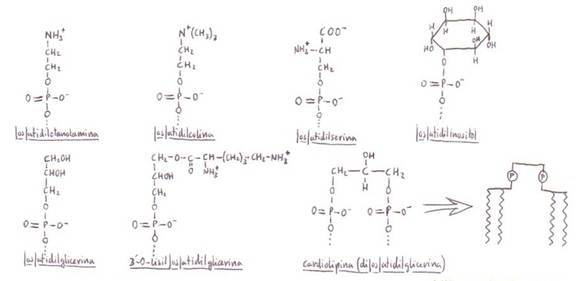
*Fosfoglicéridos (o glicerilfosfátidos): son componentes principales y
característicos de las membranas celulares.
En los fosfoglicéridos
uno de los grupos hidroxilo primarios de la glicerina
se encuentra esterificado por el ácido fosfórico; los
demás grupos hidroxilo lo están por ácidos grasos.
Además de los 2 restos de ácidos
grasos que esterifican los C1 y C2,
los fosfoglicéridos tienen un grupo de cabeza polar
que –como hemos indicado- es un alcohol (representado por X-OH) cuyo grupo
hidroxilo está esterificado por el ácido fosfórico.
Al tener una cabeza polar y
colas hirocarbonadas no polares reciben el nombre de
lípidos anfipáticos.
Habitualmente, el ácido graso en
posición 2 es insaturado y el de posición 1 saturado.
El grupo de cabeza polar de los fosfoglicéridos puede ser variado:
En plantas superiores y animales
los más abundantes son la fosfstidil-etanolamina y la fosfatidil-colina.
Son componentes principales de la mayor parte de las membranas en las células
animales.
La fosfatidil-glicerina
y la cardiolipina aparecen en membranas
celulares de bacterias. La cardiolipina,
además, en la membrana interior de la mitocondria (fue aislada en el músculo
cardiaco, donde abundan las mitocondrias).
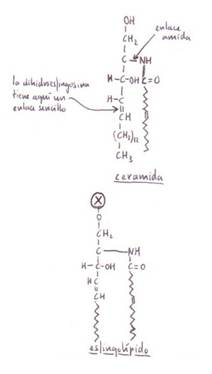
*Esfingolípidos:
su esqueleto está constituido por la esfingosina
o una base relacionada. Son muy abundantes en las membranas de células animales
y vegetales. Particularmente abundantes en el tejido nervioso y cerebral.
Todos los esfingolípidos
tienen:
-1 molécula de ácidograso.
-1 molécula de esfingosina o de uno de sus derivados.
-1 grupo de cabeza polar (en
algunos esfingolípidos
es muy complejo).
En mamíferos las bases
principales de esfingolípidos son la esfingosina y la dihidroesfingosina.
En plantas superiores y levaduras la fitoesfingosina.
La esfingosina
se une por un enlace amida a un ácido graso saturado de cadena
larga o a un monoinsaturado para dar lugar a una ceramida. Al grupo hidroxilo 1 de la ceramida se unen diversos grupos de cabeza polar.
-Esfingomielinas:
ceramida con fosforil-colina
o fosforil-etanolamina.
-Glucoesfingolípidos:
se encuentran en los centros receptores para acetil
colina y otros neurotransmisores:
.Cerebrósidos: neutros. Con un azúcar
monosacárido. En el S. N. y el cerebro son galactocerebrósidos.
.Gangliósidos: ácidos. Grupos de cabeza polisacarídicos con restos de ácido siálico.
*Ceras: son ésteres sólidos de
ácidos grasos de cadena larga con alcoholes grasos monohidroxílicos
o con esteroles (es decir: ésteres de ácidos grasos largos con alcoholes no
polares y largos). Insolubles en agua.
Cuando se calientan son blandas
y moldeables. Pero en frío son duras.
Aparecen en cubiertas
protectoras de la piel, pelo y plumas, de las hojas y frutos de plantas
superiores y en el exoesqueleto de muchos insectos.
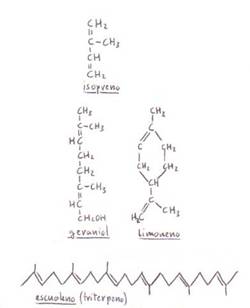
Los terpenos pueden ser lineales
o cíclicos, según la estructura que adopten.
-Monoterpenos:
geraniol
(lineal): en el aceite de geranio.
limoneno (cíclico): en el aceite de
limón.
-Sesquiterpenos:
farnesol.
-Diterpenos:
fitol (componente de la clorofila).
-Triterpenos:
escualeno (precursor de la síntesis del
colesterol).
-Tetraterpenos:
carotenoides (el β-caroteno es precursor
de la vitamina A).
*Esteroides: derivados del perhidrociclopentanofenantreno.
Todos los esteroides se originan
a partir del escualeno.
-Esteroles (con un grupo
hidroxilo en el C3):
.lanosterol:
en la cubierta cérea de la lana.
.colesterol: en la membrana
plasmática de células animales y en lipoproteínas del plasma sanguíneo.
-Ácidos biliares (con un
grupo -COOH): sintetizados en el hígado. Aperecen
unidos por el -COOH a aminoácidos. Tienen un carácter detergente (cabeza polar
y resto no polar). Ayudan en la emulsión de lípidos (para que actúen las
lipasas) y en la absorción intestinal.
-Hormonas esteroídicas:
.corticosteroides:
sintetizados en las cápsulas suprarrenales. Actúan en el metabolismo tanto
orgánico (proteínas, glúcidos y lípidos) como mineral
(reabsorción de iones en el riñón).
.hormonas sexuales: masculinas
(andrógenos) y femeninas (estrógenos y progestágenos).
*Prostaglandinas: derivados de
la ciclación de ácidos grasos insaturados de 20 C
(como el araquidónico). 5 de los átomos de C del
esqueleto carbonado del ácido graso (del C8 al C12)
forman un bucle y constituyen un anillo de 5 miembros.
Las prostaglandinas
difieren estructuralmente entre sí en el número de dobles enlaces y en los sustituyentes.
Respecto a su actividad
biológica también difieren, aunque todas presentan alguna actividad depresora
de la presión sanguínea e inductora de la contracción de los músculos lisos.