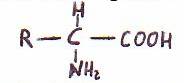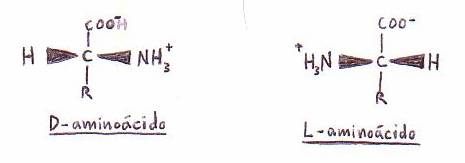Las proteínas son principios inmediatos o biomoléculas
orgánicas. Se trata de polímeros, llamados polipéptidos, formados por aminoácidos
que se unen entre sí mediante enlaces peptídicos. Al
estar formadas por aminoácidos, en su composición elemental hay -junto con C, H
y O- una elevada proporción de N. Además, puesto que existen aminoácidos proteicos
que presentan S en su estructura, el azufre también aparece en cierta cantidad.
Son las moléculas orgánicas más abundantes en los seres vivos:
constituyen aproximadamente el 50 % del peso celular en seco. Pero su importancia
no depende exclusivamente de su abundancia, sino también de la enorme variedad
de funciones que desempeñan: estructural,
de transporte de otras moléculas, movimiento, hormonal, etc. Cabe destacar su actividad como biocatalizadores (catalizadores en las reacciones metabólicas,
imprescindibles para los procesos vitales). Su función como reserva energética –aunque también se dé-
es, en cambio, menos importante: se utilizan para obtener energía cuando es
necesario debido a la falta de moléculas energéticas.
1.7.2. Aminoácidos.
Los aminoácidos constituyen el alfabeto de la estructura proteica.
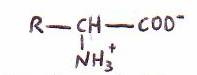
Se trata de ácidos con un grupo amino en a y con un grupo R. La
fórmula general de los 20 a-aminoácidos que se hallan corrientemente en las proteínas es:
Difieren entre sí por la estructura de sus cadenas laterales distintivas,
llamadas grupos R.
El método más significativo propuesto para clasificar los aminoácidos
se funda en la polaridad de los grupos R:
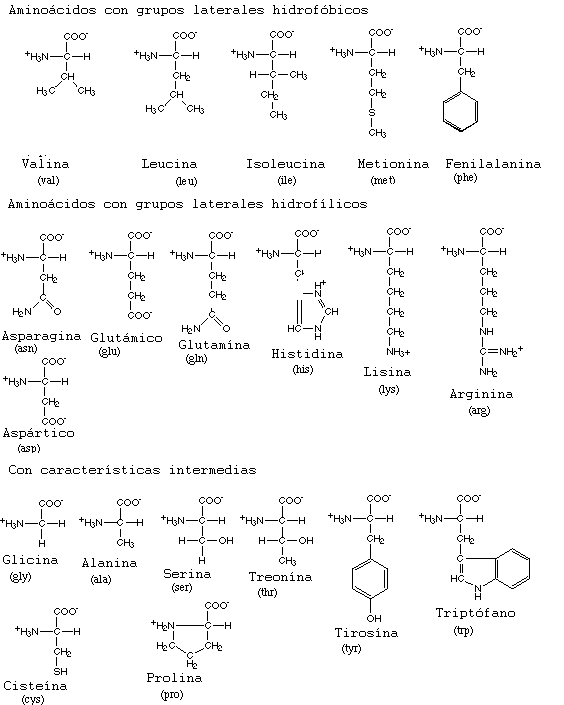
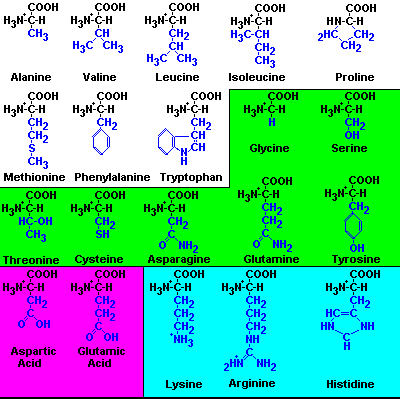
1) Aminoácidos con grupos R no polares:
-Alanina (Ala)
-Valina (Val)
-Leucina (Leu)
-Isoleucina (Ile)
-Prolina (Pro)
-Fenilalanina (Phe)
-Triptófano (Trp)
-Metionina (Met)
2) Aminoácidos con grupos R polares sin carga:
-Glicocola (Gly)
-Serina (Ser)
-Treonina (Thr)
-Cisteína (Cys)
-Tirosina (Tyr)
-Asparagina (Asn)
-Glutamina (Gln)
3) Aminoácidos ácidos (cargados negativamente a pH 6'0):
-Ácido aspártico (Asp)
-Ácido glutámico (Glu)
4) Aminoácidos básicos (cargados positivamente a pH 6'0)
-Lisina (Lys)
-Arginina (Arg)
-Histidina (His)
Además de los 20 aminoácidos corrientes en proteínas se han hallado
otros de aparición poco frecuente. Todos son derivados de algún aminoácido
corriente. Es el caso de la 4-hidroxiprolina, derivado de la Pro, que se encuentra con
cierta frecuencia en la proteína fibrosacolágeno y en algunas proteínas de las plantas. La 5-hidroxilisina
también está presente en el colágeno.
Por último, se conocen unos 150 aminoácidos más, presentes en células
y tejidos de forma libre o combinada, pero nunca en proteínas. Suelen ser
derivados de los a-aminoácidos presentes en proteínas, pero también se conocen b-, g-, y d-aminoácidos:
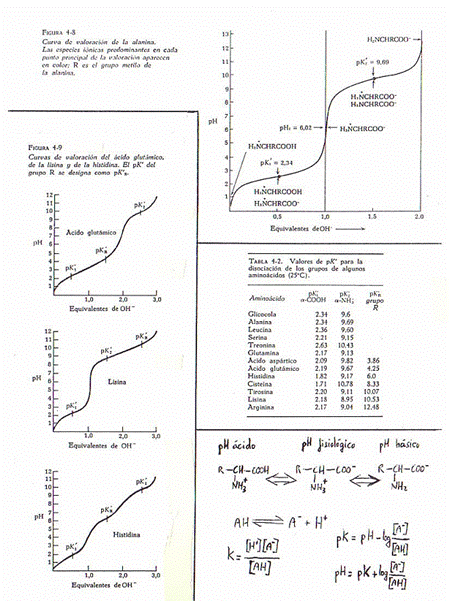
*Ácido-base: según cuál sea el pH,
los aminoácidos pueden comportarse como ácidos o como bases (es decir, como
dadores de protones o como aceptores). En medio ácido se comportan como bases y
en medio básico como ácidos, debido a que el grupo -NH2 tiende a
captar H+ en medio ácido y el -COOH a cederlos en medio básico.
Son por esto sustancias anfóteras (del gr. amphi:
ambos) y se denominan anfolitos.
A pH fisiológico aparecen así:
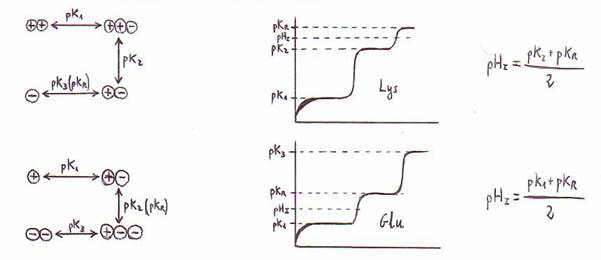
Para todos los aminoácidos hay un pH en el
que el nº de cargas + y el nº de cargas - es igual. Es el llamado pHisoeléctrico o
punto isoeléctrico.
Para aminoácidos con grupo R sin carga
pk1 + pk2
pHI = --------------
2
Para aminoácidos con grupos R cargados a pH
fisiológico
Al valorar los aminoácidos, conforme aumenta el pH
se producen por este orden los siguientes cambios:
1) a-COOH ===> a-COO-
2) x-COOH ===> x-COO-
3) a-NH3+ ====> a-NH2
4) x-NH3+ ====> x-NH2
*Actividad óptica: por ser el carbono a asimétrico, ya que
se halla unido (excepto en la Gly) a 4 radicales diferentes, los
aminoácidos tienen actividad óptica y presentan estereoisómeros:
D y L.
En las moléculas de proteínas sólo se encuentran L-aminoácidos, pero
muchos D-aminoácidos diferentes se encuentran presentes en las células.
A continuación indicamos la actividad óptica (rotación específica) de
algunos L-aminoácidos en disolución acuosa:
aminoácidorot. espec. [a]D25
L-Ala+1'8
L-Arg+12'5
L-Leu-11'0
L-Ile+12'4
L-Phe-34'5
L-Glu+12'0
L-His-38'5
L-Asp+5'0
L-Met-10'0
L-Lys+13'5
L-Ser-7'5
L-Pro-86'2
L-Thr-28'5
L-Trp-33'7
L-Val+5'6
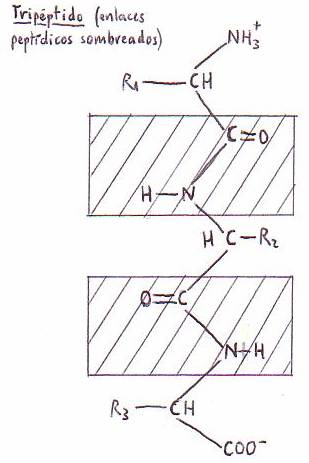
1.7.3. El enlace
peptídico. Los péptidos
a) Enlace peptídico
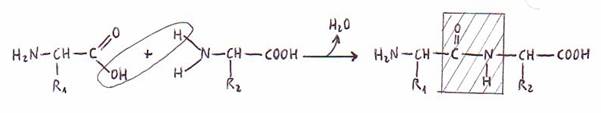
Los grupos a-amino y a-carboxilo de los aminoácidos se unen por un enlace de tipo amida que
llamamos enlace peptídico. Los péptidos pueden
llamarse, por esto, amidas sustitutivas.
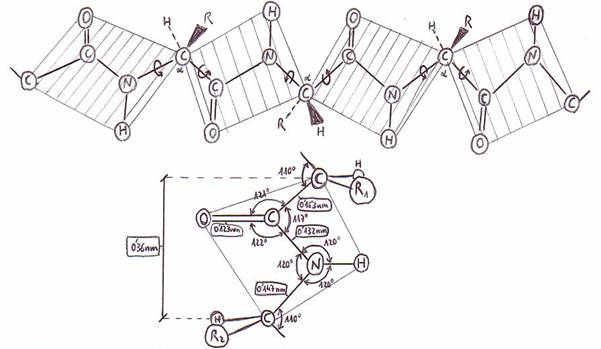
El enlace simple C-N del enlace peptídico posee alrededor de un 40% de
carácter de doble enlace, y el enlace doble C=O cerca del 40% de carácter de
enlace simple. Consecuencias:
1) El grupo imino (-NH-) del enlace
peptídico no tiene tendencia significativa a captar protones a pH fisiológico.
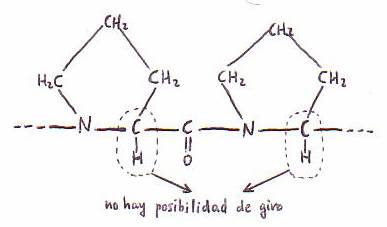
2) El enlace C-N del enlace peptídico es relativamente rígido y no
puede girar libremente (virtud importante respecto a la conformación
tridimensional de las cadenas polipeptídicas).
3) La unión C-N del enlace peptídico es más corta que la mayor
parte de los demás enlaces C-N.
El átomo de O del grupo carbonilo y el H del -NH- están en posición trans el uno respecto del otro.
Los cuatro átomos del enlace peptídico y los Ca de los aminoácidos entre los
que se establece el enlace se encuentran en un mismo plano. El único giro
posible de la cadena peptídica se da en torno a los Ca. Sólo
los enlaces sencillos de los átomos de carbono a tienen libertad de giro;
los enlaces sencillos C-N del enlace peptídico son rígidos.
El enlace peptídico impone, pues, restricciones significativas acerca
del nº de conformaciones que puede adoptar una cadena polipeptídica.
Dimensiones del enlace peptídico a partir de los datos obtenidos por
rayos X.
b) Péptidos
Cuando nos referimos a los aminoácidos, hablamos de ellos como
alfabeto de la estructura proteica. Efectivamente, los aminoácidos se combinan
entre sí -de manera similar a como lo hacen las letras del alfabeto para formar
palabras- y constituyen proteínas diversas.
Llamamos péptidos a moléculas constituidas
por aminoácidos que se unen entre sí por enlace peptídico. Según contengan 2,
3, 4 ó más aminoácidos, hablaremos de dipéptidos, tripéptidos, tetrapéptidos, etc.
Las cadenas polipeptídicas separadas o asociadas entre sí constituyen las
proteínas.
Se han identificado numerosos péptidos
cortos como productos de la hidrólisis parcial de las proteínas. También se han
encontrado en la materia viva otros muchos péptidos
no derivados de las proteínas:
*Glutatión: tripéptido
presente en los tejidos animales. Actúa como activador de determinadas enzimas
y como protector de los lípidos contra la oxidación.
Tiene en su estructura un resto de Glu unido
mediante un enlace peptídico poco frecuente en el que interviene un grupo g-carboxilo en lugar
del a-carboxilo.
*Carnosina: dipéptido del músculo.
Tiene un b-aminoácido.
*Vasopresina: nonapéptido llamado
también hormona antidiurética. Favorece la adsorción de agua y electrolitos
en el riñón.
*Oxitocina: hormona nonapeptídica que induce la contracción de la musculatura
lisa del útero.
*Antibióticos: la gramicidina
y la valinomicina.
1.7.4. Estructura de las proteínas
Como ya hemos estudiado, los a-aminoácidos se unen covalentemente entre sí mediante enlaces peptídicos y constituyen los péptidos.
Los polipéptidos (péptidos con un nº considerable de
aminoácidos) solos o asociados forman lo que denominamos proteínas.
Pero las proteínas no están organizadas sólo como una secuencia lineal
de aminoácidos, sino que dicha secuencia adopta en su estado nativo una forma
tridimensional característica que llamamos conformación. Esta
conformación tridimensional específica es necesaria para que cumplan su función
biológica específica o para que desempeñen su actividad.
Los términos que se emplean corrientemente para designar los
diferentes aspectos o niveles de la estructura proteica son: estructura
primaria, secundaria, terciaria y cuaternaria.
a) Estructura primaria
Se refiere al esqueleto covalente y establece de modo específico la
secuencia de sus restos aminoácidos. Así, denominamos estructura primaria de
una proteína a su secuencia de aminoácidos.
Los aminoácidos se encuentran unidos entre sí por enlaces peptídicos. El estudio de la estructura primaria de una
cadena polipeptídica requerirá la hidrólisis de esos enlaces y la
identificación ordenada de los aminoácidos hasta reconstruir su secuencia.
Si realizamos una hidrólisis completa de las cadenas polipeptídicas
(previa ruptura de los enlaces disulfuro
transversales por oxidación), podremos determinar la proporción que hay de cada
aminoácido pero no la secuencia.
Para determinar la secuencia lineal en laboratorio se siguen los
siguientes pasos:
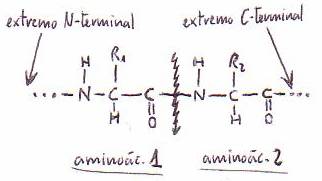
*Identificación del resto N-terminal del péptido
-Por
el método de Sanger: el dinitrofluorobenceno
(DNFB) reacciona con el N-terminal (se une y da un dinitrofenilpéptido)
-Por dansilación: el cloruro de dansilo
reacciona con el N-terminal y da un dansilpéptido.
En ambos casos a continuación se realiza la hidrólisis y se identifica
el aminoácido N-terminal.
-Por
el método de Edman: se marca el N-terminal y se separa de la cadena peptídica,
con lo que puede utilizarse por sí solo para secuenciar cadenas.
*Identificación del resto C-terminal del péptido
-Por reducción con borohidruro de litio: se transformará el grupo ácido en
un grupo alcohol. Después, cromatografía.
-Por la carboxipeptidasa: ataca el enlace peptídico COOH-terminal (hay que medir la velocidad de liberación de los
aminoácidos ya que, roto el enlace peptídico COOH-terminal,
pasa a romper el siguiente).
-Otros métodos.
*Hidrólisis parcial de cadenas
peptídicas: por enzimas específicas.
MétodoEnlaces
peptídicos escindidos
TripsinaAminoác.1: Lys o Arg
QuimotripsinaAminoác.1: Phe,
Trp o Tyr
PepsinaAminoác.1: Phe,
Trp, Tyr y otros
TermolisisnaAminoác.2:
Leu, Ile o Val
Bromuro
de cianógenoAminoác.1: Met
Mediante la combinación de estos pasos se puede deducir la secuencia
lineal y por tanto la estructura primaria del polipéptido.
b) Estructura
secundaria
Es la ordenación regular y periódica en el espacio de las cadenas
polipeptídicas a lo largo de una dirección. Se estabiliza por puentes de H
entre grupos ceto y amino del enlace peptídico.
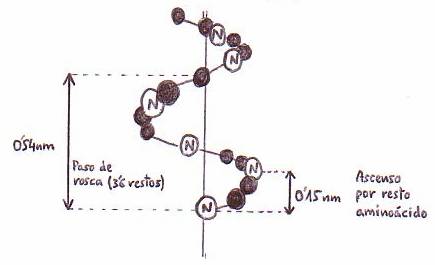
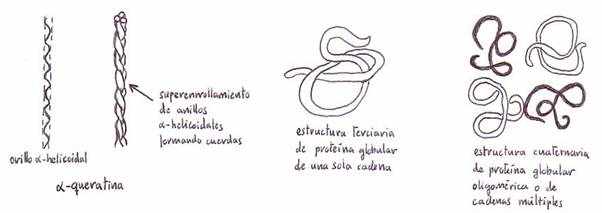
*Hélice a:
esta estructura, postulada por Pauling y Corey, tras el estudio por rayos X de las
a-queratinas, consiste en
una hélice con aprox. 3'6 restos de aminoácidos por cada vuelta. Los grupos
R
de los aminoácidos se proyectan hacia el exterior de la hélice compacta formada
por el esqueleto.
La estructura se estabiliza por puentes de H (o enlaces de H) intracatenarios.
No todos los aminoácidos forman hélice. Concretamente:
Permiten hélice a estable: Ala, Leu, Phe,
Tyr, Trp, Cys, Met, His,
Asn, Gln, Val.
Inestabilizan la hélice a: Ser, Ile, Thr, Glu, Asp, Lys,
Arg, Gly.
Rompen la hélice a: Pro, hiroxi-Pro.
Condicionantes de la estructura helicoidal (estudiados en poliaminoácidos:
polipéptidos en los que todos los restos aminoácidos son idénticos):
1) Carga del grupo R:
.poli-Ala a pH fisiológico: R pequeño y no
cargado ==> forma espontáneamente hélice a.
.poli-Glu: a pH
fisiológico: R negativo ==> no forma hélice a, pues los R se repelen.apH<2: R neutro ==> sí
forma hélice a.
.poli-Lys: a pH
fisiológico: R positivo ==> no forma hélice a.
apH>12: R neutro ==> sí
forma hélice a.
2) Volumen de R:
.poli-Ile no forma hélice a porque sus
voluminosos grupos R, próximos a los átomos Ca provocan un impedimento estérico.
3) Rigidez:
.poli-Pro y poli-HO-Pro no forman hélice a debido a que los Na
forman parte
de los rígidos anillos que constituyen el grupo
R.
Por eso siempre que aparecen la
Pro o la hidroxi-Pro en una cadena
polipeptídica, la hélice
a se interrumpe y se
origina un pliegue rígido, curvatura o codo.
La poli-Gly puede formar una hélice a, pero prefiere
otro tipo de conformación: la conformación b, en la que las cadenas
están relativamente extendidas.
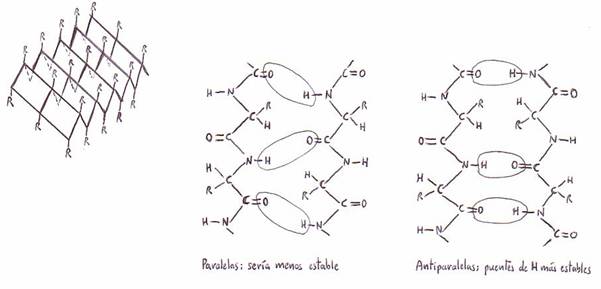
*La conformación b o en hoja plegada:
el análisis por rayos X de las b-queratinas ha revelado claves importantes para la conformación b de la cadena
polipeptídica.
Las cadenas polipeptídicas en la conformación b son paralelas y
están dispuestas en láminas plegadas que se hallan unidas
transversalmente por puentes de H intercatenarios.
Todos los enlaces peptídicos participan en
este enlace cruzado, y así confieren una gran estabilidad a la estructura. Los
grupos R se hallan por encima o por debajo de los planos en zig-zag de la lámina plegada.
Es el caso de la fibroína segregada
por el gusano de seda Bombixmori.
La fibroína y otras b-queratinas son
ricas en aminoácidos que poseen grupos R relativamente pequeños (sobre
todo Gly y Ala), mientras que los R de las a-queratinas son más
voluminosos y poseen carga mayor que los de la fibroína
de la seda.
En las formas a todas las cadenas polipeptídicas son paralelas (es decir, se
desarrollan en el mismo sentido N-terminal a C-terminal), mientras que en la fibroínalas cadenas adyacentes son antiparalelas (es
decir, se desarrollan en sentidos opuestos).
Las a-queratinas contienen muchos restos de Cys,
de forma que establecen enlaces transversales -S-S- entre cadenas
polipeptídicas adyacentes. Las b-queratinas como la fibroína no poseen
enlaces -S-S- transversales.
c) Estructura terciaria
Es la forma en la que la cadena polipeptídica se curva o se pliega
para dar lugar a la estructura globular compacta.
Influye decisivamente en la función biológica de las proteínas, ya que
ésta depende en gran medida de su estructura o forma espacial.
Para determinar y mantener esta estructura se realizan enlaces entre
diversos puntos de la cadena. Los más importantes corresponden a los puentes disulfuro entre los restos del aminoácido azufrado Cys.
Según su conformación (forma tridimensional que adoptan), las
proteínas pueden clasificarse en dos grupos principales:
*Fibrosas: formadas por cadenas polipeptídicas ordenadas
paralelamente a lo largo de un eje, formando fibras o láminas largas. Son
materiales físicamente resistentes, insolubles en agua y en las disoluciones
salinas diluidas.
Presentan función estructural en el tejido conjuntivo de los animales
superiores:
-Colágeno: en tendones y matriz de los huesos.
-a-queratina: en cabello, cuernos,
cuero, uñas y plumas.
-Elastina: en el tejido conjuntivo.
*Globulares: formadas por cadenas polipeptídicas plegadas
estrechamente, que adoptanformas
esféricas o globulares compactas. la mayor parte de
las proteínas globulares son solubles en agua.
Generalmente desempeñan una función móvil o dinámica en la célula. De
los dos millones de enzimas conocidas actualmente, casi todas son proteínas
globulares, igual que los anticuerpos, algunas hormonas y otras proteínas con función
de transporte como la seroalbúmina y la hemoglobina.
(Nota: en todas estas proteínas aparece con frecuencia un grupo prostético -de
carácter no proteico- asociado, para que desempeñen su función).
Algunas proteínas se sitúan entre los tipos fibroso y globular: se
parecen a las fibrosas por sus largas estructuras cilíndricas y a las
globulares por ser solubles en disoluciones acuosas salinas. Es el caso de la miosina
(elemento estructural del músculo) y del fibrinógeno
(precursor de la fibrina: elemento estructural de los coágulos
sanguíneos).
Una vez formada la estructura terciaria de una proteína globular, son
4 los tipos principales de interacciones o de enlaces débiles que cooperan a su
estabilización:
1) Los enlaces de H (puentes de H) entre grupos peptídicos:
como ocurre en la hélice a o en el plegamiento b.
2) Los puentes de H entre grupos.
3) Las interacciones hidrofóbicas entre
grupos no polares.
4) Los enlaces iónicos entre grupos cargados positiva y
negativamente.
d) Estructura cuaternaria
de las proteínas oligoméricas
Algunas proteínas son oligoméricas, es
decir, contienen 2 ó más subunidades polipeptídicas.
Llamamos estructura cuaternaria al modo característico mediante el las cadenas
polipeptídicas plegadas, se acoplan entre sí en la conformación nativa de una
proteína oligomérica.
Los enlaces que mantienen esta estructura son del mismo tipo que los
que mantienen la estructura terciaria.
La estructura cuaternaria de las proteínas se encuentra estrechamente
ligada a la regulación de su actividad biológica (es el caso de las enzimas alostéricas que ya estudiaremos).
Entre las proteínas oligoméricas más
sencillas está la hemoglobina (Hb), que posee
4 cadenas polipeptídicas: 2 cadenas a (141 restos cada una) y 2 cadenas b (146 aminoácidos), a cada
una de las cuales se encuentra unido un grupo hemo
mediante enlace no-covalente.
Se produce un fenómeno de unión cooperativa del O2 con la Hb:
cuanto más O2 se une a las moléculas de Hb
mayor afinidad tienen éstas por él (recordemos que en cada molécula de Hb hay 4 subunidades, cada una de
las cuales puede captar un O2 por su grupo hemo.
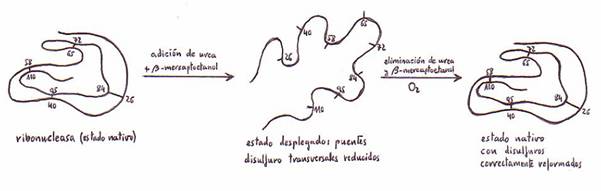
e) Desnaturalización y renaturalización de proteínas
Las proteínas globulares nativas experimentan una desnaturalización
y adoptan conformaciones desplegadas y no concretas de sus cadenas
polipeptídicas cuando se someten a:
-calefacción,
-tratamiento con ácidos o con bases,
-acción de disoluciones concentradas de urea o de cloruro de guanidina.
Con la pérdida de su conformación nativa -y en esto consiste la
desnaturalización- las proteínas pierden su actividad biológica.
Si el proceso de desnaturalización no ha sido irreversible, al
conservarse la estructura primaria, conforme retiramos los factores desnaturalizantes la proteína recupera su configuración nativa
y activa: se habla entonces de renaturalización.
(Pero en algunos casos la desnaturalización sí es irreversible).
El plegamiento se produce rápidamente. Aunque todavía no se conoce
totalmente el porqué, parece probable que este proceso se vea favorecido por el
principio de cooperatividad (intensificación
de la fuerza de atracción entre dos moléculas por la cooperación de muchos
enlaces débiles).
Algunas proteínas que carecen de enlaces disulfuro
cruzados pueden replegarse espontáneamente y con rapidez a la configuración
activa y nativa.
En el caso de las proteínas oligoméricas, cuando las sometemos a acciones que provocan
desnaturalización, experimentan cambios conformacionales
hacia formas más anárquicas en dos etapas:
1) Disociación de las cadenas
entre sí.
2) Desplegamiento
de las cadenas separadas y producción de arrollamientos al azar.
Si la desnaturalización se realiza muy cuidadosamente, puede darse el
primer paso sin el segundo. Así de ha hecho con la Hb en
presencia de sales. Al dializar el exceso de sal, las subunidades
separadas se han reasociado y la proteína ha vuelto a ser plenamente activa.
Si las condiciones son más drásticas, la desnaturalización recorre los
dos pasos. Cierto número de proteínas oligoméricas
son capaces de experimentar esta desnaturalización de forma completamente
reversible: es el caso de la enzima aldolasa
(que interviene en la glucólisis), cuyas 4 subunidades,
con la acidificación del medio se separan y despliegan, y al elevar el pH nuevamente hasta 7 se repliegan y recuperan su
estructura terciaria, y se reasocian (estructura cuaternaria) para formar la
enzima oligomérica catalíticamente activa.
1.7.5. Funciones biológicas
de las proteínas
Como sabemos, las proteínas
constituyen un grupo de biomoléculas enormemente
variado (sólo en el ser humano son decenas de miles diferentes). Ya adelantábamos
en la introducción que su importancia radica también en la gran diversidad de
funciones que desempeñan. Podemos destacar las siguientes funciones:
a)Catálisis
Hay proteínas que
intervienen como biocatalizadores (catalizadores de las reacciones biológicas),
y forman parte de las enzimas. Estas
moléculas favorecen las reacciones químicas en los seres vivos incrementando su
velocidad. Serán objeto de estudio en la siguiente unidad. Dentro de este grupo
podemos citar la catalasa, la ribonucleasa,
las isomerasas, deshidrogenasas,
etc.
b) Transporte
Algunas proteínas se
unen a diversas sustancias para transportarlas con mayor eficacia por el medio
interno. Cabe destacar aquí la hemoglobina
(que transporta el oxígeno en la sangre) y las lipoproteínas (que transportan lípidos). Dentro de las lipoproteínas plasmáticas se distinguen:
quilomicrones,
que transportan triacilglicéridos y colesterol exógenos
(de la dieta) desde el intestino hasta el hígado; lipoproteínas de muy baja densidad (VLDL) y de baja densidad (LDL), que llevan triacilglicéridos
y colesterol endógenos (sintetizados por el organismo) desde el hígado hacia
los tejidos (el llamado vulgarmente “colesterol malo”); y lipoproteínas de alta densidad (HDL), que transpostan
colesterol desde los tejidos al hígado (el llamado “colesterol bueno”).
c) Inmunidad
Existen proteínas que
proporcionan la identidad molecular de los organismos vivos y (antígenos), y otras que rechazan –anulándola-
cualquier molécula extraña que se introduzca en el organismo (anticuerpos). Los anticuerpos son inmunoglobulinas (moléculas formadas por
proteínas y glúcidos), que estudiaremos con
detenimiento al final del temario.
d) Homeostasis
Muchas
proteínas actúan directamente para mantener la estabilidad del medio interno
(recordemos que el conjunto de procesos encaminados a este fin era llamado homeostasis): algunas, por su capacidad
amortiguadora, intervienen en el mantenimiento del pH;
la trombina y el fibrinógeno actúan en la
coagulación de la sangre, y evitan pérdidas en el medio interno; algunas
hormonas intervienen en la circulación de la sangre para mantener constante la
temperatura del cuerpo; etc.
e) Función hormonal
Hay hormonas de
naturaleza proteica que intervienen en el metabolismo de glúcidos
(como la insulina y el glucagón), y
otras que participan en procesos de desarrollo (como la somatotropa u hormona del crecimiento).
f) Función estructural
Aquí se incluyen proteínas
que forman parte de estructuras celulares (como las glucoproteínas de la membrana
plasmática, las proteínas del citoesqueleto celular y
de los flagelos y cilios, y las histonas que mantienen la estructura de la cromatina). También
el colágeno y la elastina de tendones y ligamentos, y la queratina de uñas y pelos.
g) Contracción muscular
La miosina y la actina
son filamentos proteicos que constituyen las miofibrillas
responsables de la contracción muscular.